Epithelial tissue grows through a combination of cell division (mitosis), collective cell migration, and differentiation, all tightly orchestrated by signaling pathways, physical architecture, and mechanical cues. In short: stem cells divide in specific zones, daughter cells migrate outward or upward, and the tissue expands as a coordinated sheet, not as a pile of independent cells. Once you understand those three levers, you can start predicting what goes wrong when growth stalls and what conditions push it forward.
How Does Epithelial Tissue Grow? Key Mechanisms and Checks

Marcus Whitmore
29 May 2026
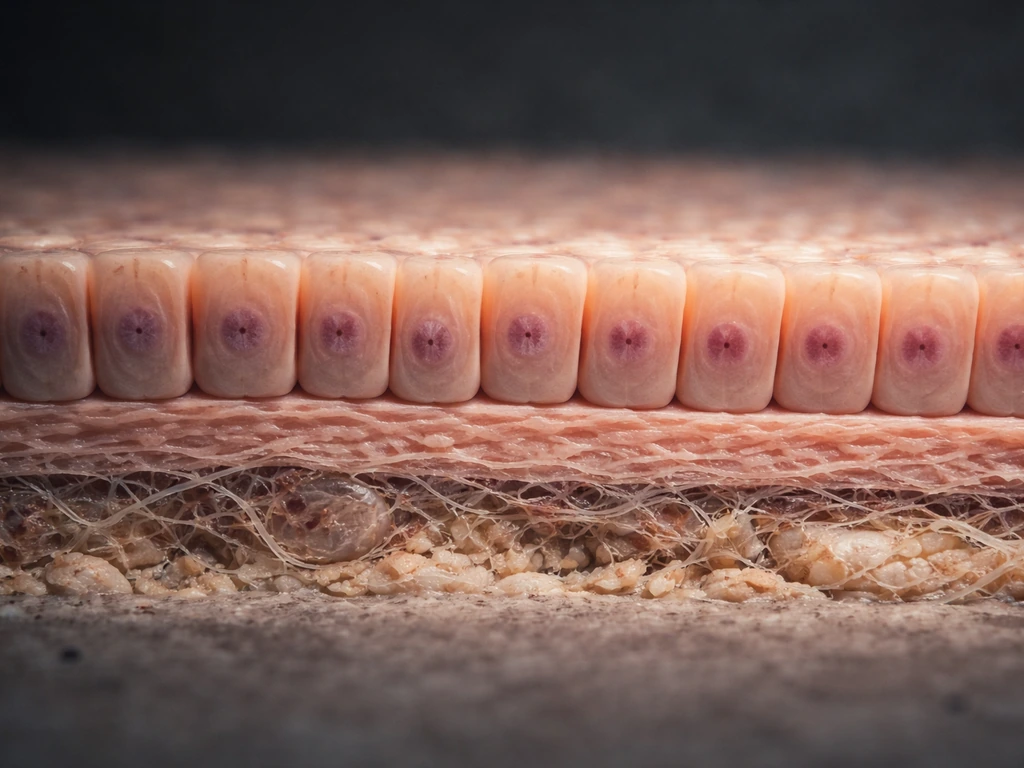
What epithelial tissue is and where it actually grows

Epithelial tissue is the body's lining material. It covers every surface, inside and out: the outer layer of skin, the gut lining, the airways, blood vessel walls, kidney tubules, and gland surfaces. If something separates your body from the outside world, or separates one compartment of your body from another, it's almost certainly epithelium.
All epithelial tissue rests on a basement membrane, a non-cellular mat of proteins and carbohydrates (like collagen IV and laminin) secreted by both epithelial and underlying connective tissue cells. That basement membrane isn't just structural padding. It's an active platform that anchors the basal layer, supplies growth signals, and acts as a physical boundary that epithelial cells must respect. Growth that violates this boundary, like when cancer cells breach the basement membrane, is a sign that normal control has broken down.
The two best-studied sites of continuous epithelial growth in adults are the epidermis (skin) and the intestinal lining. Both maintain a clear spatial logic: stem cells sit in a protected zone, divide, and push progeny outward. In skin, stem cells occupy the basal layer and progeny move up toward the surface. In the intestinal lining, stem cells sit at the very bottom of crypts, and daughter cells migrate upward to the villus tip, where they're shed into the gut lumen every two to six days. Knowing where growth originates in a given epithelium tells you almost everything about how to study or stimulate it.
The three main mechanisms: division, migration, and differentiation
Cell division: the engine of expansion
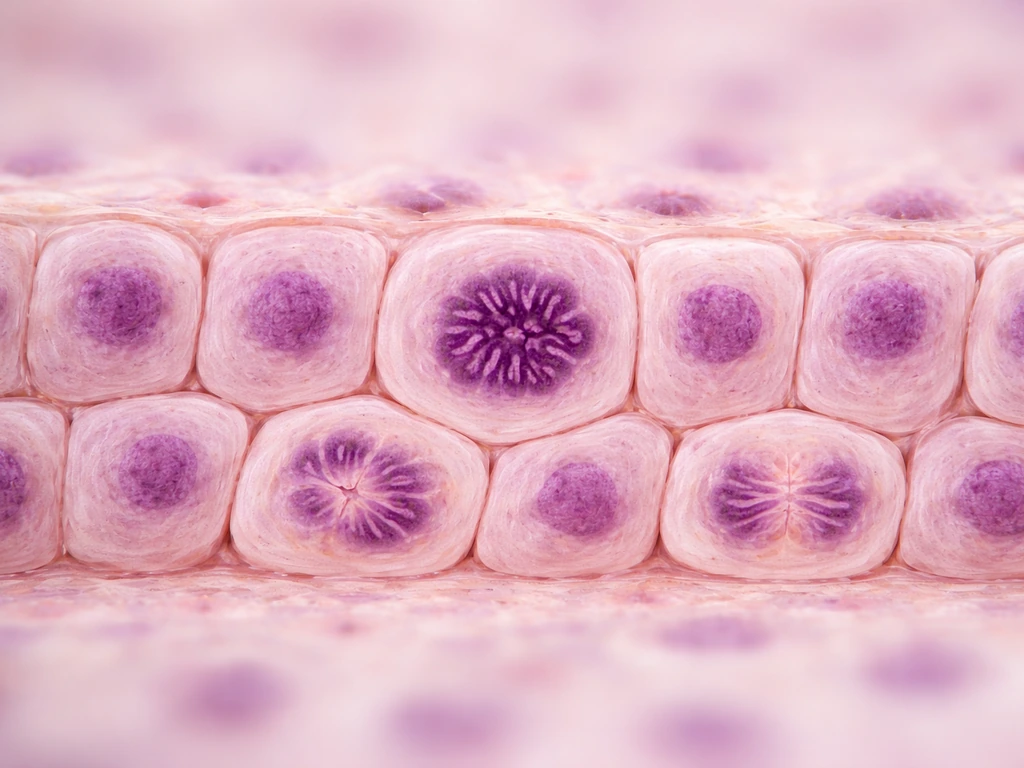
Mitosis is the obvious mechanism, but where and when it happens matters enormously. In the epidermis, proliferative keratinocyte stem cells (capable of unlimited self-renewal) and transit-amplifying (TA) cells coexist in the basal layer. TA cells divide a limited number of times before their daughters withdraw from the cell cycle entirely and commit to differentiation. In the intestine, Lgr5-expressing stem cells at crypt position +1 do the dividing, with daughter cells progressively differentiating as they migrate up. Proliferation is regionally restricted, not spread uniformly across the tissue. If you're looking for where growth happens in any epithelium, start by asking where the stem cells are.
Collective migration: moving as a team
Here's the part most people underestimate: epithelial growth is often driven by migration before it's driven by division. This is most obvious in wound healing. Within hours of injury, existing keratinocytes at the wound edge dissolve their hemidesmosomes (the protein complexes anchoring them to the basement membrane), reorganize their cytoskeleton, and slide into the defect as a coherent sheet. Proliferation becomes the dominant theme only a few days later. A small incisional wound can be fully re-epithelialized within 24 to 48 hours in humans, largely through migration alone. For larger wounds, epithelial cells advance at roughly 1 to 1.5 mm per 10 days in limb wounds, until the migrating edges meet.
This migration isn't random. Epithelial sheets move collectively, with 'leader cells' at the front activating EGFR (epidermal growth factor receptor) signaling and coordinating the movement of the cells behind them. If you block E-cadherin, the glue that keeps cells connected to each other, the coordinated sheet falls apart and wound closure becomes irregular. The collective nature of epithelial migration is both its strength and its constraint: the tissue can cover ground fast, but only if cell-cell contacts are intact.
Differentiation: growing without multiplying

Not all epithelial expansion comes from new cells. Differentiation, the process by which a dividing progenitor becomes a specialized functional cell, is the step that converts raw proliferation into a working tissue layer. In the skin, dividing basal cells give rise to layers of keratinocytes that flatten, fill with keratin, and eventually die to form the protective outer surface. In the intestine, stem-cell progeny differentiate into absorptive enterocytes, mucus-secreting goblet cells, hormone-secreting enteroendocrine cells, and antimicrobial-peptide-releasing Paneth cells. A tissue that can only divide but not differentiate won't function. Both processes are required for true growth.
The control panel: signaling pathways, growth factors, and the cell cycle
Think of the signaling environment as the control panel that decides when division happens, when it stops, and when progenitors switch to differentiation. In the intestine, three pathways do the heavy lifting: Wnt, Notch, and BMP. Wnt and Notch together maintain the intestinal stem cell pool at the crypt base. Remove either signal and the stem cells are lost. Wnt also drives proliferation by activating β-catenin/TCF transcriptional targets including c-Myc and cyclin D1, which push cells into the cell cycle. Mechanistically, Wnt-driven intestinal hyperproliferation requires the cyclin D2-CDK4/6 complex, directly linking pathway activation to G1/S cell-cycle progression.
In the epidermis, EGF and its receptor are central players. The clinical relevance here is direct: topical epidermal growth factor has been tested in randomized controlled trials to accelerate epidermal regeneration at skin graft donor sites, and it works. This connects basic biology to a practical intervention you can actually look up and evaluate. Growth factors don't just tell cells to divide; they also regulate migration and survival. EGF, for example, drives both mitosis and keratinocyte motility.
The Hippo pathway, specifically the transcriptional co-activators YAP and TAZ, sits at the intersection of mechanical signals and growth control. When cells are under high mechanical tension (on a stiff substrate, or in a stretched tissue), YAP/TAZ moves into the nucleus and drives proliferative gene expression. When cells are soft, rounded, or in contact with many neighbors, YAP/TAZ is held in the cytoplasm and growth is suppressed. Knocking down YAP/TAZ impairs wound healing and reduces proliferation. This means that even the physical stiffness of the environment is part of the growth control system, not just the chemical signals.
How tissue architecture shapes growth

Architecture isn't just anatomy. The spatial organization of an epithelial tissue actively regulates whether cells grow, stop, or migrate. Four structural features are especially important.
- Basement membrane: anchors basal cells via integrins (especially α6β4 integrin binding laminin-332), provides growth factor depots, and defines the basal boundary. Stable hemidesmosomal adhesion to the basement membrane is a 'brake' on migration; releasing it (by disassembling hemidesmosomes) is one of the first steps in wound re-epithelialization.
- Apical-basal polarity: epithelial cells have distinct apical (lumen-facing), lateral (cell-cell contact), and basal (substrate-facing) membrane domains. Polarity is established and maintained by tight junctions, adherens junctions, and polarity complexes. Loss of polarity is closely linked to loss of growth control.
- Tight junctions: these occluding junctions block passage between adjacent cells, maintaining barrier function and also acting as upstream sensors for the Hippo pathway. Tight junctions and adherens junctions both feed into YAP/TAZ regulation, connecting barrier integrity directly to growth suppression.
- Contact inhibition: when epithelial cells form a complete monolayer and touch their neighbors on all sides, proliferation stops. This contact inhibition of growth is mediated largely through the Hippo pathway. An open wound edge removes that constraint, which is exactly why wound-edge cells start moving and then dividing.
An elegant demonstration of how architecture drives growth comes from organoid experiments. A single Lgr5-expressing intestinal stem cell, placed in the right extracellular matrix and growth factor conditions, can self-organize to build full crypt-villus structures in vitro without any mesenchymal support cells. The cell doesn't need a pre-made blueprint handed to it from the outside; the stem cell itself reads its local signals and recreates the architecture. That's a profound point: the tissue growth program is intrinsic, but it requires the right physical and chemical context to execute.
Physical constraints: why epithelial growth has limits
Epithelial tissue can't grow indefinitely, and the reasons are partly biological and partly physics. Understanding these limits helps you interpret when growth is genuinely constrained versus when it's just receiving the wrong signals.
| Constraint | What it limits | Practical implication |
|---|---|---|
| Space and geometry | Available surface area for sheet expansion; wound geometry determines migration distance | Incisional wounds (<1 mm gap) close in 24-48 hours; large excisional wounds take days to weeks |
| Nutrient and oxygen supply | Proliferating cells have high metabolic demand; diffusion limits how far cells can be from supply | Thick tissues require vascularization; epithelial sheets are thin for good reason |
| Contact inhibition | Cells stop dividing once they form a confluent, fully connected sheet | Complete epithelial coverage restores growth suppression via Hippo/YAP-TAZ |
| Basement membrane integrity | Migration requires either an intact or provisional matrix substrate; absence of substrate stalls migration | In chronic wounds, degraded ECM can block re-epithelialization |
| Stem cell pool size | Growth rate is capped by the number of dividing stem and transit-amplifying cells available | Exhausting or damaging the stem cell pool (e.g., radiation) causes permanent epithelial failure |
Tissue tension is worth calling out separately. As a wound closes, the mechanical tension in the epithelial sheet increases. That rising tension both provides a growth signal (via YAP/TAZ activation) early on and eventually contributes to signaling that the gap is closed. The wound geometry itself, its depth, area, and the condition of the surrounding tissue, determines the rate at which cells can migrate and whether proliferation needs to catch up. This is why wound shape matters clinically: a circular punch wound closes differently than a linear incision.
Regeneration vs development: wound healing, stem cells, and normal turnover
There's an important distinction between how epithelial tissue grows during development and how it regenerates after injury. During embryonic development, epithelial sheets expand rapidly over large areas, using a mix of proliferation and coordinated migration that is more permissive than in the adult. Adult wound repair recapitulates some, but not all, of that developmental program. Adult wound re-epithelialization shows dramatically increased proliferation that parallels embryonic epidermis formation, but the regulatory environment is tighter and the signals more redundant. The authors report that adult wound repair shows dramatically increased proliferation during re-epithelialization that parallels aspects of embryonic development after migration initiation Building vs. Rebuilding Epidermis.
Normal tissue turnover sits between those two extremes. The intestinal epithelium is one of the fastest-renewing tissues in the body: the entire functional villus epithelium is replaced every two to six days, driven continuously by crypt stem cells. Protein turnover rates across intestinal segments run at roughly 0.16 to 0.20 per day, which means the lining is essentially rebuilt multiple times per week. The epidermis turns over more slowly but follows the same logic: stem cells divide, progeny differentiate, surface cells are shed. This constant renewal is why radiation or chemotherapy, which preferentially kills dividing cells, often damages gut and skin epithelium first.
Wound healing adds a burst of growth on top of that baseline. The four phases (hemostasis, inflammation, proliferation, remodeling) overlap in time, but for epithelial growth, the key window is the transition from inflammation to proliferation, roughly days one through three after injury. Keratinocytes begin migrating within hours. By around day three, epithelial stem cells in the basal layer and hair follicle root sheaths begin proliferating to supply the advancing front. Once re-epithelialization is complete, contact inhibition and restored cell-cell contacts shut the proliferative signal down again. If that shutdown fails, you get hypertrophic scarring or, in extreme cases, malignant transformation.
What to do with this today: practical guidance for real-world epithelial growth problems
If you're working with epithelial growth right now, whether in a wound care context, in cell culture, or as a student designing an experiment, here's the most direct advice I can give you.
If growth isn't happening, check these things first
- Is the substrate right? Epithelial cells need a basement-membrane-like matrix (laminin, collagen IV, fibronectin) to anchor and migrate on. In culture, use Matrigel or appropriate ECM coatings. In a wound, check whether the wound bed has viable matrix or is covered in necrotic tissue blocking migration.
- Are the growth factors present? EGF, keratinocyte growth factor (KGF/FGF7), and hepatocyte growth factor (HGF) are the main drivers of epithelial proliferation and migration. Serum-free media must supplement these deliberately. In a chronic wound, these signals may be degraded by excessive proteases.
- Is the mechanical environment appropriate? Cells on very soft substrates suppress YAP/TAZ and reduce proliferation. If you're culturing on an overly compliant surface, or if a wound is under zero tension, growth signals are dampened. Gentle mechanical stimulation or appropriate substrate stiffness matters.
- Are cell-cell contacts intact but not over-confluent? A sparse monolayer with gaps grows toward confluence. A fully confluent monolayer stops growing by contact inhibition. If you want to study active growth, seed at ~30-50% confluency. If wound closure has stalled in a model, check whether cells have gone fully quiescent.
- Is the stem cell pool intact? In a severe injury or after cytotoxic treatment, the stem cells themselves may be depleted. No amount of growth factor supplementation will fix an absent stem cell pool. This is the scenario where cell-based therapies (stem cell transplants, skin grafts) become necessary.
What to measure and observe
In any epithelial growth experiment, track at least three things: the rate of sheet expansion (scratch assay closure rate or wound planimetry), the proliferation index (Ki67 or EdU staining of dividing cells in the basal layer or crypt), and cell morphology/polarity (look for the cobblestone appearance of healthy epithelium versus elongated, mesenchymal-looking cells that have undergone epithelial-to-mesenchymal transition). YAP localization (nuclear versus cytoplasmic) is an excellent readout of mechanical status and growth state if you have immunofluorescence available.
Connecting the biology to the clinical or experimental context
In a clinical wound care setting, the practical levers are: debridement to remove matrix barriers to migration, moisture-retentive dressings to maintain the hydrated environment cells need to migrate, and, in selected cases, topical growth factor therapy (EGF-based preparations have clinical trial evidence behind them for donor site wounds). In an organoid or in vitro model, the levers are matrix composition, growth factor cocktail, and seeding density. In a developmental biology context, the questions shift to spatial patterning of stem cell zones and gradient-driven signaling, but the same three mechanisms (division, migration, differentiation) still apply.
One final point worth remembering: epithelial growth and the broader question of how cells grow and divide are deeply connected topics. The same cell-cycle machinery (cyclin-CDK complexes, checkpoint proteins) that governs how any cell grows also controls epithelial expansion, just with tissue-specific regulatory layers built on top. If you're studying how cells grow or increase in size more broadly, the epithelial system is one of the most instructive models you can use, because the spatial organization makes the growth logic visible. In general, cell growth in size is driven by the balance between cell-cycle progression, nutrient availability, and signals from the surrounding environment cells grow or increase in size. In many tissues, though, the most noticeable changes during growth often reflect coordinated proliferation and differentiation rather than cells permanently getting bigger as they grow do cells get bigger as you grow. Adipose tissue growth follows similar principles, with region-specific stem or progenitor activity, coordinated expansion, and context-dependent signaling that governs when cells proliferate and when they differentiate. To understand why do cells grow, it helps to view growth as the outcome of coordinated division, migration, differentiation, and signaling control why cells grow.
Quick-reference takeaways
- Epithelial tissue grows from regionally restricted stem cell zones, not uniformly across the tissue surface.
- Migration comes before proliferation in wound healing; don't assume 'growth' always means 'dividing.'
- The basement membrane is both anchor and signal platform; damaged or absent matrix blocks re-epithelialization.
- Contact inhibition is a feature, not a bug. When it fails, growth becomes pathological.
- Mechanical cues (substrate stiffness, tissue tension) are real growth regulators via YAP/TAZ, not just background noise.
- If growth has stalled, check substrate, growth factors, mechanical environment, confluency, and stem cell viability, in that order.
- Wnt, Notch, BMP, and EGF/EGFR are the main pathway families driving epithelial proliferation and differentiation across tissues.
FAQ
If epithelial cells are proliferating, why might the tissue not actually grow or close a wound?
Epithelial expansion is not governed by one switch. In intact tissue, division tends to be restricted to defined stem zones, while the “output” of that division depends on whether cells can migrate, maintain adhesion, and complete differentiation. If you block cell-cell adhesion (for example, via E-cadherin disruption), you can still see individual proliferation, but the tissue sheet will not close smoothly.
How does epithelial regrowth differ between small vs large wounds, especially early on?
Yes, but the growth speed and success depend on whether migration can physically span the gap while the cells remain polarized and attached to a suitable substrate. Small wounds often re-epithelialize largely by migration first, whereas larger defects may require that the migrating front is continually supplied by new progenitors once the distance exceeds what existing cells can cover.
What happens to epithelial regeneration if the stem-cell “maintenance” signals are reduced?
Loss of stem-cell maintenance signals typically reduces the size of the renewing cell pool at the tissue base, which then causes a delayed failure of regeneration even if migration and differentiation are initially intact. In other words, you may see temporary repair, but turnover cannot be sustained once the stem reservoir is depleted.
Can epithelial tissue expand without proper differentiation, and what would that look like functionally?
Differentiation can be “necessary” even when the tissue is expanding. If progenitors keep dividing but cannot enter the correct differentiation programs, you get a layer that may cover the surface but lacks proper function, polarity, and barrier properties, and it may remain in a hyperproliferative state.
What tells epithelial cells to stop proliferating after a wound is closed?
Contact inhibition is a key stop signal once the epithelial sheet restores coverage. Practically, this means proliferation typically drops after the wound closes because cell-cell contacts and tissue architecture are re-established. If cells fail to re-establish proper contacts, proliferation can persist and contribute to abnormal thickening.
How does basement membrane integrity influence normal growth versus invasive behavior?
Basement membrane integrity provides anchoring cues and a boundary that normal epithelium respects. If cells cannot properly attach to or traverse that boundary, migration and organization fail; if they breach it without the usual controls, you move toward invasive, dysregulated growth behavior rather than normal renewal.
What are common experimental reasons epithelial scratch assays look like they require proliferation when they really don’t?
In culture, sheet closure rates are strongly affected by how dense you seed and how well cells form a confluent, polarized monolayer before you scratch. Low density or poor confluence makes migration appear slower and can make it seem like “division is required” when the real limitation is loss of collective behavior.
How should I interpret nuclear YAP or TAZ localization when studying epithelial growth under different mechanical conditions?
YAP and TAZ activity is a mechanotransduction readout, but it is not a simple linear “more stiffness, more growth” relationship across all contexts. To interpret it, you need to consider substrate stiffness, cell density, and whether the tissue is stretched or constrained, because these variables change both mechanical tension and cell-cell contact patterns.
Which measurements best distinguish migration-driven epithelial closure from proliferation-driven expansion?
Cell-cycle markers like Ki67 or EdU show proliferation, but they do not distinguish whether growth is driven by migration, division, or both. Pairing proliferation staining with geometric closure metrics (wound planimetry or sheet-edge tracking) helps separate “cells dividing” from “sheet advancing.”
Why do organoids sometimes fail to build normal crypt-villus structures even when stem cells are present?
Organoid self-organization depends on providing the right extracellular matrix and growth factor context, but also on correct starting geometry and exposure timing. If matrix composition or growth factor gradients are off, you can get partial structures or abnormal patterning even when cells are viable.
Why do chemotherapy or radiation often damage intestinal and skin epithelia first, and what determines recovery?
Because epithelial turnover is continuous, interventions that primarily target cycling cells can cause disproportionately rapid epithelial injury. That means “recovery” depends on how quickly the stem zone can restore the proliferative pool and how intact the basement membrane and architecture remain.
Next Article
How Do Cells Grow or Increase in Size? Key Mechanisms
How cells grow and increase size via cell cycle, biosynthesis, membrane biogenesis, and division, limited by transport a