After a pollen grain lands on the stigma, it germinates and sends a tube growing down through the style toward the ovule. That tube does not wander randomly. It is pulled and pushed in a very specific direction by a chain of chemical signals, physical tissue channels, and molecular machinery operating at the tube's growing tip. Water absorption triggers the whole process, a calcium gradient steers it at the cellular level, and peptide attractants secreted by cells near the egg literally lure the tube to its destination. The style is not a passive pipe but an active guide, and every stage of that journey is constrained by biology, chemistry, and the physical limits of the tissue.
How the Pollen Tube Grows Down the Style Step by Step

Marcus Whitmore
30 May 2026
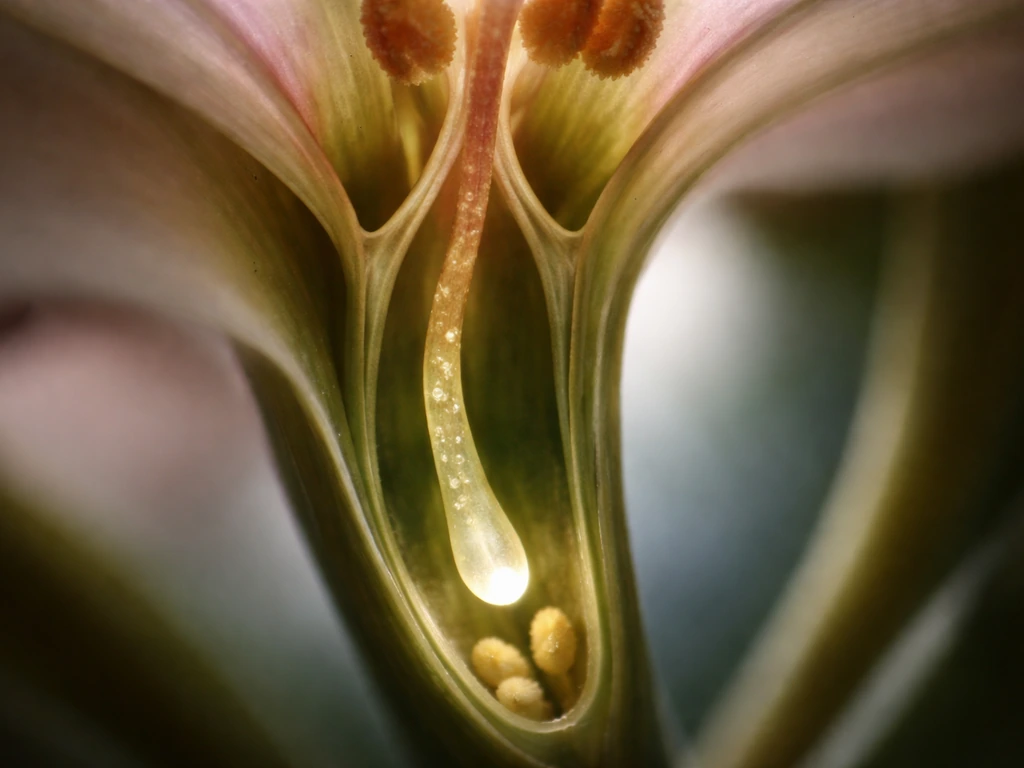
What triggers pollen tube growth after pollination

The moment a compatible pollen grain lands on a receptive stigma, it starts absorbing water. That hydration is not just a passive soak. It rapidly builds turgor pressure inside the grain, and that internal pressure is the physical engine that pops the tube out through a thin region of the pollen wall called the aperture. Think of it like squeezing a water balloon through a small opening: the pressure finds the weak point and the tube begins to emerge.
Extracellular calcium ions play an essential role right from this first step. Without available Ca2+ in the surrounding environment, germination either fails or stalls. Once the tube starts growing, calcium becomes even more important: a steep tip-focused Ca2+ gradient builds up inside the tube, with the highest concentration right at the growing tip. This gradient is not incidental. It is essential. Channels at the tip, including cyclic nucleotide-gated channels (CNGCs) and stretch-activated Ca2+ channels, continuously let Ca2+ flow in, maintaining that gradient and keeping growth directed and active.
At the same time, the pollen grain's own energy reserves, primarily sugars like sucrose, fuel the biosynthesis and vesicle trafficking needed to build new membrane and wall material. Research on tobacco pollen tubes shows that depleting sucrose dramatically reduces tube elongation, which tells you the grain is not just being pushed by turgor alone. It is also actively manufacturing the growth machinery as it goes.
This is also where self-incompatibility comes in. If the pollen and style share matching S alleles (in gametophytic self-incompatibility systems), the style produces S-RNases that enter the pollen tube and destroy RNA, halting growth. In Brassica-type (sporophytic) self-incompatibility, a stigma receptor kinase (SRK) recognizes the pollen's own protein ligand, triggers a phosphorylation cascade, and blocks the very hydration step before a tube can even form. The germination trigger is real, but so is the shutdown mechanism that catches self-pollen early.
How the style guides the tube: signals and tissue structure
The style is not just a stretch of plant tissue the pollen tube drills through blindly. It has a specialized internal structure called the transmitting tract, a channel lined with secretory cells that produce an extracellular matrix rich in proteins, polysaccharides, and signaling molecules. This tract is the highway, and it is built to be navigated. The tube follows the transmitting tract down toward the ovary, guided both by the physical channel and by the chemical environment the tract creates.
Guidance works in stages. In the early part of the journey through the style, the transmitting tract tissue and its surface-expressed or secreted cues do most of the directing. Think of this like a river channel: the tube tends to follow the path of least resistance and highest chemical attractiveness through the stylar tissue. As the tube exits the style and approaches the ovule, a different guidance system takes over. Cells called synergids, located right next to the egg cell inside the ovule, secrete small cysteine-rich peptides called LUREs. These diffuse outward through the micropyle (the tiny opening in the ovule's outer layers) and create a concentration gradient the pollen tube tracks directly toward.
The molecular basis for this LURE attraction is now well established. In Arabidopsis, a receptor on the pollen tube surface called AtPRK6 binds directly to AtLURE1 peptides secreted by the synergid cells. This receptor-ligand interaction is species-specific, which is part of why cross-species fertilization usually fails: the attractant and receptor do not fit together. So the guidance system is not generic diffusion toward any chemical gradient. It is a lock-and-key system that ensures species-appropriate fertilization.
Mechanical factors matter too. The physical stiffness of the stylar tissue and the geometry of the transmitting tract actively shape which direction the tube can grow. Evidence suggests guidance is not purely chemical. The pistil tissue creates mechanical constraints that reinforce the directional signal. Chemical cues and mechanical walls work together to keep the tube on track. The eLife article on pollen tube guidance emphasizes that directed growth is shaped by both chemical guidance cues and mechanical cues from pistil and stylar tissues Chemical cues and mechanical walls work together to keep the tube on track..
How the pollen tube grows at the tip: membrane, wall, and vesicle delivery
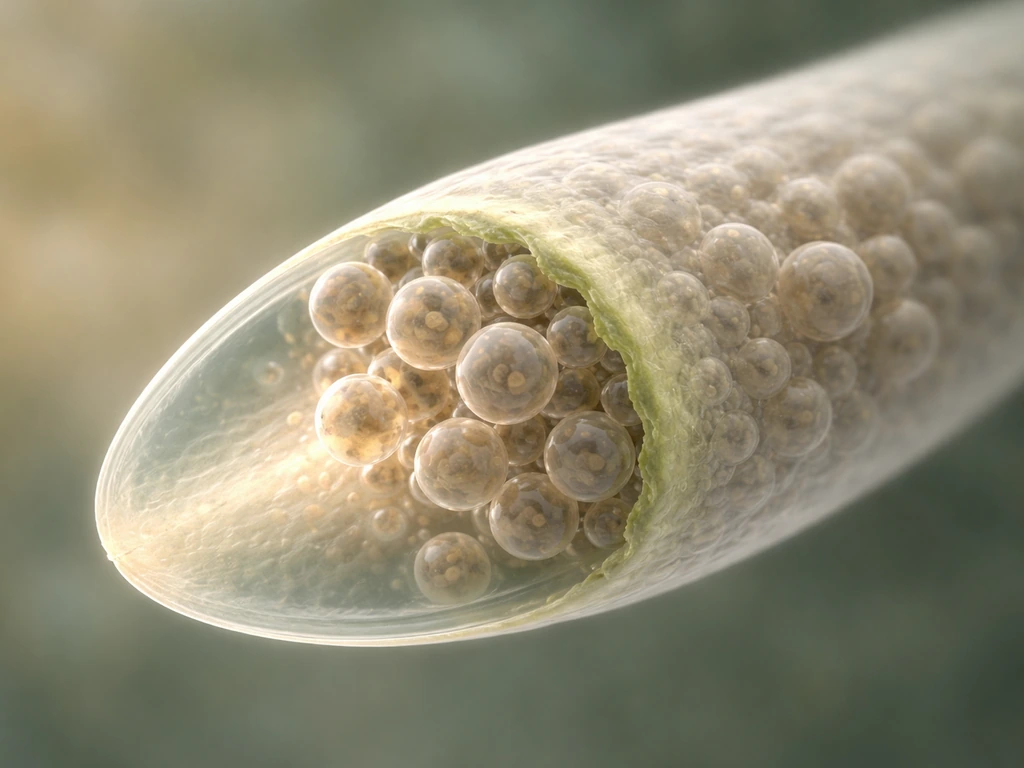
Here is where growth mechanics get genuinely fascinating. The pollen tube only grows at its very tip. The rest of the tube, the shank behind it, is essentially locked in place with a hardened cell wall. All the extension action happens in a tiny apical domain a few micrometers across. Understanding this is key to understanding everything else about pollen tube growth.
What drives tip extension is a continuous delivery of Golgi-derived vesicles to the apex. These vesicles carry cell wall material (mainly pectins) and membrane lipids. When they fuse with the plasma membrane at the tip (exocytosis), they deposit fresh, soft wall material and expand the membrane. Turgor pressure then pushes the newly deposited soft wall outward, extending the tube. It is a tight collaboration: vesicle delivery brings the raw material, turgor provides the force. Neither alone is enough.
The cytoskeleton is the logistics system running this delivery. Actin filaments (F-actin) form a dynamic network at the tip, organized by a small signaling protein called ROP1 GTPase. ROP1 activates downstream pathways (RIC4 and RIC3) that promote actin assembly and disassembly in coordinated bursts. This oscillating actin organization drives oscillatory vesicle delivery, which is why pollen tube growth is not a smooth, steady extension but a rhythmic, pulsing process. You can actually measure growth rate oscillations in real time in the lab.
The cell wall chemistry at the tip is carefully managed. Freshly delivered pectins are methylesterified, meaning they are in a soft, plastic form that can deform under turgor. Behind the tip, an enzyme called pectin methylesterase (PME) removes methyl groups from the pectin. Demethylesterified pectin then crosslinks with Ca2+ ions, stiffening the wall. So the tip stays soft enough for growth while the shank firms up to resist turgor blowout. PME inhibitor proteins (PMEIs) at the apex prevent premature hardening right where growth needs to happen. The whole setup is basically a self-reinforcing system: growth at the front, consolidation at the back.
Callose (a beta-1,3-glucan polymer) adds another layer to this design. As the tube extends, callose plugs form periodically behind the growing tip, partitioning the old cytoplasm from the active growing zone. These plugs are not random deposits. They act like one-way valves, preventing backward flow and keeping the metabolically active cytoplasm concentrated near the tip where it is needed. Staining with aniline blue or calcofluor white highlights these callose deposits beautifully under fluorescence microscopy, which is how researchers track tube growth in plant tissue in vivo.
Why it grows down (directionality) instead of randomly: constraints and feedback
This is probably the question that trips people up most. Why does the tube grow consistently toward the ovule rather than spiraling in circles or reversing? The answer is that directionality is enforced at multiple levels simultaneously, not by a single magic signal.
At the molecular level, the tip-focused Ca2+ gradient is the primary compass. Ca2+ influx is highest at whichever part of the tip membrane is currently extending. If a guidance cue (like a LURE peptide gradient) nudges higher Ca2+ influx slightly to one side, the actin network and vesicle delivery shift toward that side, bending growth in that direction. The ROP1 signaling module at the apex is exquisitely sensitive to spatial asymmetries in these signals. This is very similar in principle to how other tip-growing cells (like root hairs and fungal hyphae) steer: a local bias in signaling at the tip gets amplified into a directional growth response.
At the tissue level, the transmitting tract physically channels the tube. The extracellular matrix of the tract is not uniform. It has composition gradients and surface cues that preferentially support tube adhesion and penetration along the stylar axis. The tube cannot easily grow sideways through the dense stylar tissue wall; the path of least resistance and maximum chemical support is downward.
The callose plug system reinforces this too. By sealing off the tube behind the tip, callose prevents any reversal of cytoplasmic flow. Growth becomes a one-way process structurally, not just chemically. Combined with the external LURE gradient near the ovule pulling the tip forward, and the transmitting tract channeling growth through the style, you end up with a robust, multiply redundant directional system. This same logic of growth guided by evolving signals echoes the larger idea of what weaves webs as they grow in nature. If one guidance mechanism is perturbed in experiments, the others often compensate partially, which is why it has taken researchers a long time to unpack each individual layer.
This multi-layered directionality is a great example of a general principle worth recognizing: directional growth in biology almost never depends on a single cue. Whether you are thinking about how microtubules grow toward specific cellular targets, how a coleoptile bends toward light, or how a pollen tube tracks an ovule, you tend to find layered, redundant mechanisms ensuring that growth goes the right way. Microtubules in many cells behave similarly, alternating between polymerizing growth and shrinkage to drive directional tip growth microtubules grow toward specific cellular targets. Like a pollen tube, a coleoptile’s growth direction and rate can be predicted from the light-driven signals it experiences and how they shape cell expansion coleoptile bends toward light. The pollen tube is just an unusually clear and well-studied example.
What conditions affect growth rate and success

Growth rate varies enormously between species and between in vivo and in vitro conditions. In vitro, Zea mays (corn) pollen tubes have been measured growing at around 1.14 micrometers per second, while Ornithogalum virens tubes grow at roughly 0.25 micrometers per second. Some Conospermum species show some of the fastest recorded rates. These are not just fun facts: the variation tells you that growth rate is not a fixed biological constant but a reflection of the tube's interaction with its specific chemical and physical environment.
The key environmental variables that control growth rate and success are:
- Calcium availability: External Ca2+ is required to maintain the tip gradient. Too little and the gradient collapses, growth stalls or the tube bursts.
- Sugar supply: Sucrose in the medium (or provided by style secretions in vivo) fuels vesicle production and osmotic balance. Depleting sucrose in tobacco pollen tube experiments dramatically reduces elongation.
- pH: Pollen tube growth rate oscillates alongside cytoplasmic pH changes. The apical region has a measurably different pH from the shank, and disrupting this gradient impairs growth.
- Boric acid: Boron is a co-factor in pectin crosslinking in the cell wall and is required for normal germination and tube growth in vitro germination media.
- Temperature: Like most enzymatic and membrane processes, pollen tube growth has an optimal temperature range and slows significantly outside it.
- Stylar compatibility: If incompatibility signals are triggered (as in self-incompatibility), growth is actively halted in the style regardless of all other conditions being favorable.
- Osmolarity of the medium or surrounding tissue: Incorrect osmolarity causes either tube bursting (too low) or plasmolysis and growth arrest (too high).
In vitro pollen tube growth assays are very sensitive to medium formulation. Small differences in how pollen is dried onto agar, or in the concentration of calcium salts and sucrose, can produce wildly different germination percentages and tube lengths. This is why comparing in vitro growth rates across studies requires caution and why researchers developing apple cultivar assessments, for example, emphasize standardizing incubation conditions carefully. What you observe in a petri dish is real biology, but it reflects the environment you created, not necessarily what happens inside a flower.
Common misconceptions and how to visualize the process effectively
The biggest misconception is that the pollen tube is like a drill that physically forces its way through the style by brute mechanical pressure alone. It is not. The tube grows through the transmitting tract because the tract is designed to allow and guide it, and because enzymatic secretions from the tube tip help remodel the extracellular matrix ahead. Turgor is essential but it works in coordination with a softened, receptive tissue. Imagine not a drill through concrete but a root tip pushing through moist soil: the environment cooperates.
Another common sticking point is confusing the pollen grain with the pollen tube. The grain is a multicellular structure sitting on the stigma. The tube is a new cellular extension that emerges from the grain and carries two sperm cells (in angiosperms) to the ovule. The grain itself does not travel anywhere. Students sometimes picture the whole grain migrating down the style, which is not what happens.
People also tend to think of the growing tip as a rigid structure being pushed from behind. In reality the tip is the softest, most dynamic part of the entire tube. The shank behind is the stiff, callose-reinforced section. Growth happens only at the front, and the chemistry of the wall is deliberately kept plastic there to allow it.
To build a good mental model, try visualizing it in three stages: first, a water-swollen grain sitting on the stigma, with a bubble of new membrane forming at the aperture under turgor pressure. Second, that bubble extending into a narrow tube that threads into the transmitting tract, with vesicles constantly arriving at the tip and depositing new wall material as turgor pushes the front forward. Third, that tube homing in on the micropyle of an ovule, following a chemical gradient of LURE peptides like a guided wire.
If you want to observe this directly, aniline blue staining under UV fluorescence is the practical go-to technique. Callose in the tube walls and plugs glows bright blue-white, letting you see exactly how far tubes have grown through whole pistil sections. Paired with time-lapse microscopy in vitro, you can watch the oscillatory growth pulses in real time and see the tip zone clearly. That visual experience fixes the concept in a way that diagrams alone rarely do.
Finally, it is worth stepping back and recognizing why this process is such a clean illustration of constrained, directional growth. The pollen tube cannot grow faster than its vesicle supply allows. It cannot grow sideways against the transmitting tract architecture. It cannot grow past a self-incompatible style. Every limit is a specific biological constraint, not a vague ceiling. That pattern, where growth rate and direction are both tightly coupled to the environment and tissue structure the growing structure inhabits, shows up across biology from microtubule dynamics to how root tips navigate soil. Microtubules in many cells grow in similarly constrained, directional ways, with local signaling and mechanics shaping where growth occurs microtubule dynamics. The pollen tube is one of the most tractable systems we have for studying it.
FAQ
What is the first signal that tells a pollen grain to start growing a pollen tube down the style?
The earliest trigger is hydration on the stigma. Water uptake causes the grain to build internal turgor pressure and rapidly reorganize growth machinery at the aperture, then calcium availability and tip-focused Ca2+ gradients sustain the tube once extension begins.
How does the pollen tube know which direction is “down” when the style’s shape can vary?
Directionality is enforced by more than one system. The transmitting tract provides a physical, tissue-lined path that favors downward penetration, while tip Ca2+ asymmetry (steered by cue gradients such as LURE near the ovule) biases vesicle delivery to the correct side.
If Ca2+ gradients steer the tip, what happens if Ca2+ levels are low or uneven?
Low extracellular Ca2+ can prevent germination or stall early tube growth. Even after growth starts, disrupting Ca2+ gradients reduces the accuracy of tip steering because actin organization and vesicle traffic become less spatially biased.
Does the pollen tube grow only because it is being pushed, or does it actively build new material as it extends?
It actively builds. Turgor supplies the force, but elongation requires continuous delivery of Golgi-derived vesicles to the apex, which add soft wall components and expand the membrane. Without vesicle supply, pressure alone cannot extend the tube.
Why does the pollen tube not grow everywhere along its length, only at the tip?
The shank is typically stabilized by hardened, callose-reinforced cell walls, while the very apex remains a plastic growth zone. The wall-chemistry switch (plastic at the tip, stiffening behind) limits expansion to a few micrometers at the front.
How do callose plugs prevent the tube from reversing or leaking cytoplasm?
Callose plugs form behind the active tip and partition older cytoplasm from the growing zone. They act like barriers that reduce backflow, so the system remains effectively one-way, even if conditions fluctuate.
What is the role of ROP1 and actin in directing growth down the style?
ROP1 at the apex organizes an oscillating actin network. When signaling becomes slightly asymmetric, actin rearrangements shift where vesicles land at the tip, turning small directional biases into sustained bending and forward extension.
Why do species often fail to fertilize across borders, even if a pollen tube reaches the ovule?
LURE attraction is species-specific at the receptor level. If the ovule’s peptide cues do not match the pollen tube’s receptor, the guidance signal will not “lock in,” so the tube may not be properly attracted through the micropyle.
How does self-incompatibility stop a pollen tube, and at what point can it act?
Depending on the system, it can block the earliest hydration step (so the tube never starts) or degrade tube RNA after germination. In both cases the outcome is growth arrest, because the style deploys recognition mechanisms against matching S alleles.
Can enzyme secretion from the pollen tube actually change the style environment?
Yes. While the transmitting tract architecture channels growth, tube tip activity can also remodel extracellular matrix ahead of the tip. This reduces mechanical resistance locally, helping the tube penetrate through tissue that is not just a simple empty pipe.
What are common mistakes when visualizing this process in models or diagrams?
Two frequent errors are assuming the whole pollen grain travels down the style (it does not) and imagining the tube as a rigid drill pushed from behind. The grain stays put, and only the tip is soft and actively extending, with the rest becoming progressively reinforced.
Why can pollen tube growth rates look different between in vivo and lab assays?
Because the tube’s interaction with its environment changes. Media composition, especially calcium salts and sucrose, and differences in how pollen is prepared can alter germination percentage, growth speed, and tube length, so cross-study rate comparisons need careful standardization.
How can I confirm where a tube has grown when working with stained tissue sections?
A practical approach is callose-selective staining, since callose deposits highlight the tube and its plugs. Observing fluorescence under appropriate imaging conditions lets you measure penetration distance through pistil sections rather than guessing from a single time point.
Next Article
Can Microtubules Grow From Both Ends? Plus vs Minus Growth
Learn how microtubules have plus and minus ends with asymmetric growth, dynamic instability, and factors that affect eac
