Mitochondria and chloroplasts are the two organelles that can genuinely grow and divide to reproduce themselves. They do this through a process of binary fission, much like bacteria, and they carry their own DNA to make it possible. A handful of other organelles, including peroxisomes, can also multiply by growth and division, but they work differently. Most other organelles, like the endoplasmic reticulum (ER), Golgi, and nucleus, are simply distributed intact to daughter cells when a cell divides rather than being built from scratch or truly self-replicating.
Which Organelles Can Grow and Divide to Reproduce

Marcus Whitmore
31 May 2026
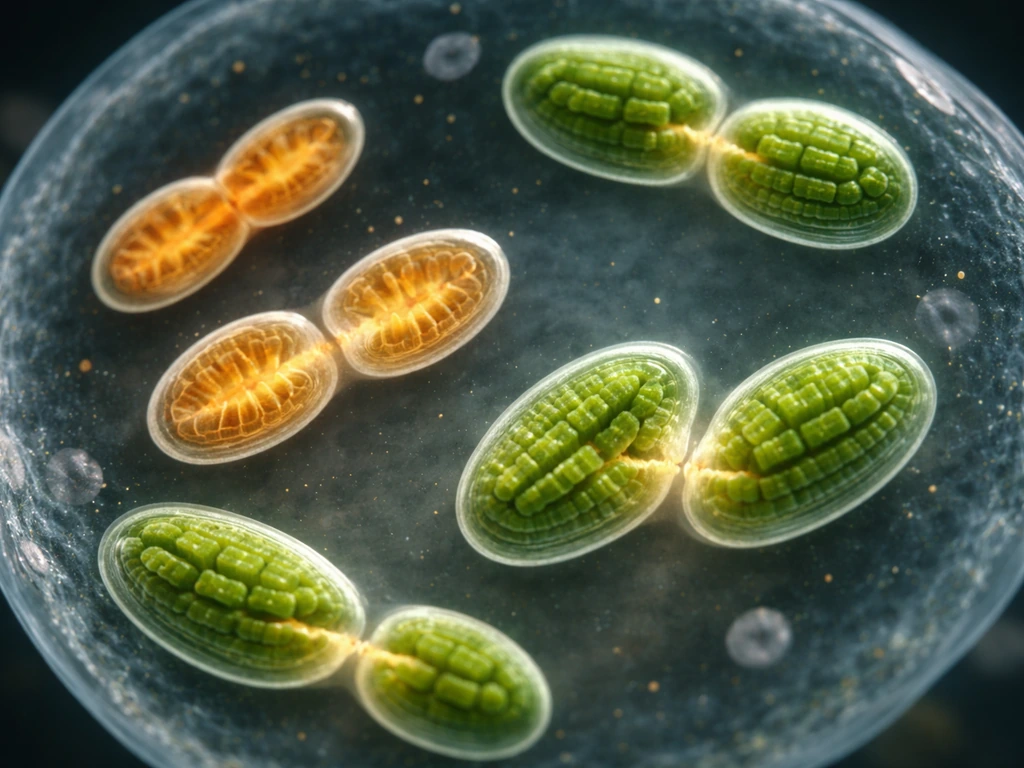
Which organelles can actually self-replicate?

The short list of self-replicating organelles falls into two categories: those with their own genomes and those without. Mitochondria and chloroplasts sit in the first category. Both descended from free-living bacteria that were engulfed by ancestral cells over a billion years ago, and both retained a stripped-down version of a bacterial genome. That retained DNA is a critical part of what lets them divide independently of being built from scratch each time a cell needs more of them.
Peroxisomes sit in the second category. They have no genome at all, yet they can multiply by growth and division. They are sometimes described as 'self-replicating' organelles in textbooks, but that label is a bit misleading: peroxisomes depend entirely on imported proteins encoded by the nucleus, and under some conditions they can even be generated de novo from the ER. Mitochondria and chloroplasts, by contrast, cannot be regenerated de novo. If you experimentally eliminate all mitochondria from a cell, the cell cannot rebuild them from nothing. That irreversibility is a defining feature of truly autonomous organelles.
Every other common organelle, including the ER, Golgi, lysosomes, and the nucleus itself, relies on being inherited intact when a cell divides. They are not assembled fresh and they do not divide autonomously. The information needed to build them is not encoded in the genome alone, it is also embedded in the physical structure of the existing organelle.
How organelles grow in size and then divide
Mitochondria: fission driven by DRP1
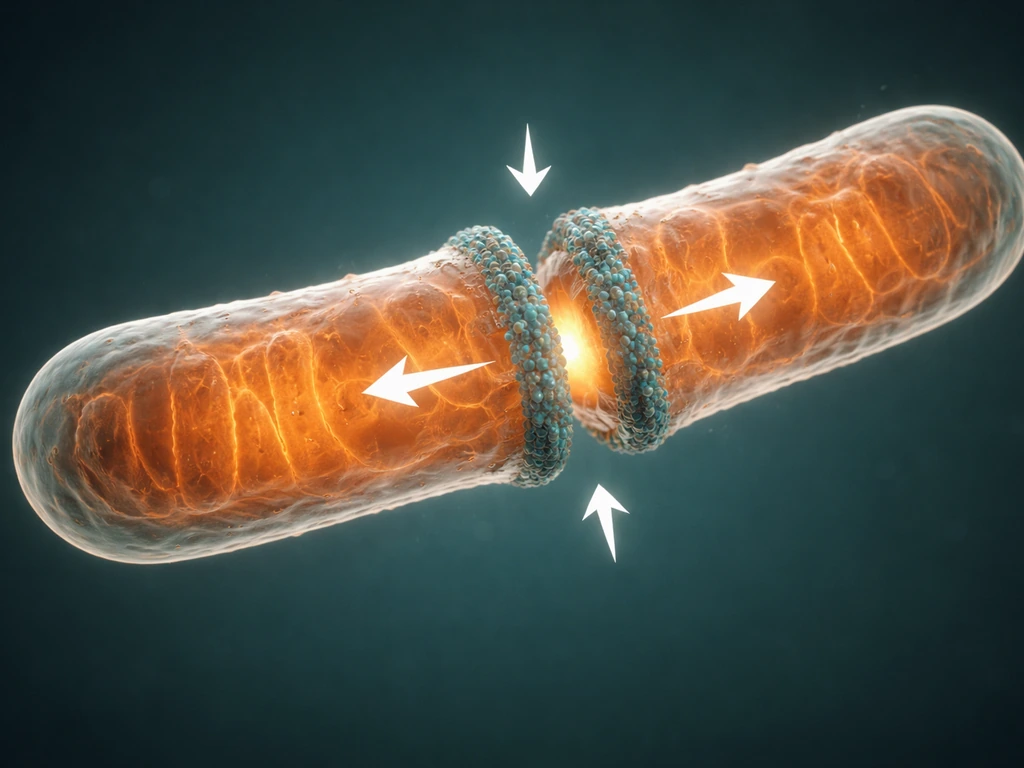
Mitochondrial growth starts with biogenesis: the organelle increases its membrane mass and protein content before it pinches apart. Most of the roughly 1,500 proteins that make up a mitochondrion are encoded in the nucleus, synthesized in the cytoplasm, and imported through two translocase complexes. The TOM complex (translocase of the outer membrane) handles proteins crossing the outer membrane, while TIM23 (translocase of the inner membrane) ferries matrix-destined proteins inward. That import pipeline is not optional; without a steady flow of nuclear-encoded proteins, a mitochondrion cannot grow or divide.
Once a mitochondrion has grown large enough, fission begins at sites where the ER wraps around the mitochondrion and physically constricts it. That ER contact site is also where mtDNA replication machinery, specifically the polymerase POLG2, tends to cluster, meaning the cell couples DNA replication with division in the same physical location. The actual scission is driven by DRP1 (dynamin-related protein 1), a GTPase that is recruited from the cytoplasm to the outer mitochondrial membrane by adapter proteins including Mff and MiD49/MiD51, as well as Fis1. DRP1 assembles into a ring around the constriction point and uses GTP hydrolysis to mechanically pinch the membrane until the mitochondrion splits in two.
DRP1 activity is tightly regulated by post-translational modifications: phosphorylation, ubiquitylation, SUMOylation, and S-nitrosylation all shift its behavior. This is how the cell keeps mitochondrial division from running out of control. It is not enough for DRP1 to simply be present; it has to be in the right activation state, at the right location, at the right time in the cell cycle.
Chloroplasts: the Z-ring and the dynamin ring
Chloroplast division is more elaborate and uses a machinery that is unmistakably bacterial in origin. The process requires the coordinated assembly of four protein rings spanning both envelope membranes. Inside the chloroplast, FtsZ (a tubulin-related protein inherited from the cyanobacterial ancestor) polymerizes into a Z-ring at the inner envelope membrane. The inner-envelope protein ARC6 stabilizes that ring. On the cytoplasmic side of the outer envelope, PDV1 and PDV2 recruit the dynamin-related GTPase ARC5 (also called DRP5B) to form an outer ring that mirrors the inner one. The two rings constrict in concert, and the chloroplast is pinched apart.
Like mitochondria, chloroplasts must grow before they divide: new membrane lipids are synthesized, chloroplast-encoded proteins are made by the organelle's own ribosomes, and nuclear-encoded proteins are imported through the TOC and TIC translocon systems (the chloroplast equivalents of TOM and TIM). Protein import into chloroplasts occurs post-translationally across the chloroplast envelopes using the TOC and TIC translocons (outer- and inner-envelope translocases, respectively) nuclear-encoded proteins are imported through the TOC and TIC translocon systems. Key TOC receptors include Toc159 and Toc34, with Toc75 forming the channel. Without that import traffic, biogenesis stalls.
What conditions have to be in place for organelle replication
Think of organelle reproduction as having a checklist. All of the following need to be in place before a mitochondrion or chloroplast can successfully grow and divide: This is similar to how organelles have a specific set of prerequisites before they can replicate within human cells before human cells can grow and reproduce they need.
- Membrane raw materials: phospholipids and other lipids must be available to expand the organelle's membranes as it grows.
- Protein supply: nucleus-encoded proteins must be continuously synthesized in the cytoplasm and imported through TOM/TIM (mitochondria) or TOC/TIC (chloroplasts).
- DNA replication: the organelle's own genome must be replicated before division. In mitochondria this requires POLG (DNA polymerase gamma), TWINKLE helicase, mtSSB, and TFAM. In chloroplasts, analogous prokaryotic-type replication factors operate.
- Division machinery activation: DRP1 (mitochondria) or FtsZ/ARC5 (chloroplasts) must be recruited and activated in a spatially and temporally controlled manner.
- Energy availability: ATP is needed for every step, from import motors to GTPase-driven constriction.
- Cell cycle permissiveness: the host cell's regulatory signals must allow or promote organelle division, typically timed with or just before the cell's own division.
For chloroplasts specifically, light is also a factor. Studies on spinach leaf discs show that light drives chloroplast DNA synthesis and times the chloroplast division cycle. And in Chlamydomonas, chloroplast DNA replication is regulated by the redox state of the organelle independently of division itself, meaning the organelle is biochemically monitoring its own metabolic health before committing to replication.
Energy, materials, and staying in sync with the rest of the cell
Organelle self-replication is metabolically expensive, and the resources do not come from the organelle alone. Human cells also need plenty of energy and raw materials, plus tight regulatory control, to successfully grow and reproduce. Mitochondria produce ATP through oxidative phosphorylation, but they cannot make all of their own membrane lipids or synthesize most of their own proteins. They are dependent on the rest of the cell for both. In return, mitochondria supply the ATP that the cell needs to run protein synthesis and import, which feeds back into mitochondrial biogenesis. It is a mutual dependency that makes organelle reproduction genuinely cooperative rather than autonomous.
TFAM (mitochondrial transcription factor A) is a good example of how the cell keeps a hand on the throttle. TFAM binds directly to mtDNA, both packaging it into nucleoids and regulating transcription and replication from mtDNA promoters. The amount of TFAM in a mitochondrion is tightly controlled by nuclear gene expression. If you want more mitochondria, you need more TFAM. If TFAM levels drop, replication slows. That nuclear control of a mitochondrial factor is a clean illustration of how 'self-replicating' these organelles really are: they replicate under the supervision of the nucleus.
For chloroplasts, the communication runs in both directions through a process called retrograde signaling. When chloroplast redox state is disturbed, reactive oxygen species (ROS) and redox signals travel back to the nucleus and alter nuclear gene expression. This feedback loop adjusts the rate at which nuclear-encoded chloroplast proteins are produced, which in turn throttles biogenesis and division. The chloroplast is essentially reporting its own condition back to the control center and waiting for permission to proceed.
Why organelle replication is not unlimited
You might wonder: if mitochondria and chloroplasts can divide, what stops a cell from filling up with them? Several hard limits exist, and they are worth understanding because they connect directly to broader principles of how size is constrained in living systems.
- Physical space: a cell has a fixed volume. As organelles multiply, they compete for cytoplasmic space, and a cell that is too packed cannot function.
- Membrane surface-to-volume limits: building more organelle membrane requires more lipid, which has to be synthesized by the cell. Lipid synthesis capacity is finite.
- Protein import throughput: every nucleus-encoded mitochondrial or chloroplast protein has to be imported through a finite number of translocon complexes. That import rate sets a ceiling on how fast organelle mass can increase.
- DRP1 and FtsZ availability: division machinery proteins are present in limited pools. If DRP1 is all recruited and occupied, new fission events cannot be initiated without synthesis of more.
- Regulatory control via post-translational modifications: as described above, DRP1 is switched on or off by phosphorylation and other modifications. These act as brakes on runaway division.
- Host cell cycle gating: organelle division is loosely coupled to the cell cycle. Signals that promote cell division also tend to promote organelle division, but the cell cycle itself has checkpoints and limits.
This connects to a pattern you see across biology: growth at every level, from organelles to whole organisms, is not limited by a simple on/off switch but by a convergence of resource availability, physical constraints, and regulatory feedback. The same logic that explains why cells cannot grow indefinitely (surface-to-volume ratio problems, transport limits, DNA replication timing) applies at the sub-cellular scale too. These same trade-offs also determine how cells grow and reproduce to maintain homeostasis, rather than expanding without limit.
Mitochondria vs. chloroplasts vs. peroxisomes: a quick comparison
| Feature | Mitochondria | Chloroplasts | Peroxisomes |
|---|---|---|---|
| Own genome? | Yes (mtDNA, ~16.5 kb in humans) | Yes (cpDNA, ~120–160 kb in plants) | No |
| Division mechanism | DRP1-mediated fission at ER contact sites | FtsZ Z-ring + ARC5/DRP5B dynamin ring | Growth and division; some de novo from ER |
| Division machinery origin | Eukaryotic dynamin-related | Bacterial FtsZ + eukaryotic dynamin-related | Mixed; PEX proteins |
| Protein import system | TOM/TIM translocases | TOC/TIC translocases | PEX receptors/importins |
| Can be regenerated de novo? | No | No | Yes (under some conditions) |
| Key regulatory factor | DRP1, TFAM | FtsZ, ARC6, PDV1/2 | PEX11 |
| Division coupled to cell cycle? | Yes, via DRP1 modifications | Yes, via FtsZ expression levels | Partially |
Real-world examples: what 'grow and divide' actually looks like
Mitochondria in muscle cells during exercise training
When you train aerobically, your muscle cells respond by increasing mitochondrial mass, a process called mitochondrial biogenesis. This happens through nuclear signaling (via PGC-1alpha and downstream pathways), which upregulates TFAM expression, boosts protein import capacity, increases the rate of mtDNA replication, and promotes DRP1-driven fission to distribute the new mitochondrial mass into more, smaller mitochondria. None of this happens because the mitochondria 'decided' to multiply; it happens because the nucleus received signals from physical activity and adjusted the conditions that allow mitochondrial growth and division to proceed.
Chloroplasts dividing in a developing leaf
In a young leaf just emerging from a bud, cells start with a small number of proplastids (immature, undifferentiated plastids). As the leaf expands and is exposed to light, those proplastids differentiate into chloroplasts and begin dividing rapidly. FtsZ expression increases, the Z-ring assembles, and binary fission produces dozens of chloroplasts per cell. In a mature spinach leaf cell, you might find 50 to 100 chloroplasts, each one the descendant of a proplastid. Crucially, none of this division could happen without the nucleus supplying the proteins and the cell providing the energy and membrane materials.
What organelles cannot do
Even the most 'self-sufficient' organelle cannot be isolated from the cell and made to divide in a test tube without the full complement of host-supplied proteins, membranes, and energy. Purified mitochondria in vitro lose their ability to divide within hours. No organelle, not even mitochondria or chloroplasts, can be generated from scratch if the starting population is eliminated. That is a fundamental constraint that sets organelle reproduction apart from the reproduction of free-living cells.
Yeast cells, unlike these organelles, are full free-living organisms that follow their own cell cycle to grow, then reproduce by budding or dividing how yeast cells grow and reproduce. Think of it this way: a bacterium is a complete self-replicating unit; a mitochondrion is a highly streamlined fragment of one, permanently dependent on its host.
How to use this knowledge practically
If you are working through a biology course or trying to answer an exam question about which organelles can grow and divide to reproduce themselves, here is how to think through it clearly:
- Start with the endosymbiotic organelles: mitochondria and chloroplasts are the primary answer. They have their own genomes, they divide by binary fission, and they cannot be regenerated de novo.
- Add peroxisomes as a secondary case: they can multiply by growth and division too, but they lack a genome, can be reformed from the ER, and work by entirely different mechanisms (PEX proteins rather than FtsZ or DRP1).
- Distinguish 'self-replicating' from 'truly autonomous': no organelle is fully autonomous. All depend on the host cell for protein import, lipids, energy, and regulatory signals. 'Self-replicating' means they grow and divide using inherited templates, not that they work in isolation.
- Connect the mechanism to the conditions: for any organelle to divide, it needs raw materials, import-competent translocons, active division machinery, a replicated genome (if applicable), and appropriate cell cycle signals.
- Trace the constraints: physical space limits, import throughput, regulated division machinery, and host cell gating all prevent unlimited organelle proliferation, mirroring the constraints that prevent unlimited cell or organism growth.
If you want to go deeper on related mechanisms, it is worth looking at how cells themselves grow and reproduce, what conditions they need to divide, and how those requirements scale up to whole-organism growth. Understanding organelle replication is really understanding a miniaturized version of the same principles that govern everything from dividing yeast cells to the growth of an entire organism: materials must be available, energy must flow, and regulatory checkpoints must be passed before growth can proceed. Similarly, stem cells also need the right materials, energy, and regulatory checkpoints to make cell division possible, which is the same core idea behind what do stem cells need to grow and divide. For yeast cells, growth and reproduction also rely on coordinated cell-cycle control, nutrient supply, and cell division machinery dividing yeast cells.
FAQ
Are peroxisomes “self-replicating” in the same way mitochondria and chloroplasts are?
Besides mitochondria and chloroplasts, peroxisomes can multiply, but they are not autonomous. Their proteins come from the nucleus, and they can form de novo under some conditions by budding from the ER, so they do not behave like mitochondria or chloroplasts whose genomes are organelle-intrinsic and whose replication cannot be rebuilt from nothing.
What happens if the organelle protein import system is blocked, but the organelle’s own DNA replication machinery is intact?
Mitochondria and chloroplasts can divide only if there is a functioning import system for nuclear-encoded proteins. In practice this means you can get growth signals and even some DNA replication, but division still fails if the TOM/TIM (mitochondria) or TOC/TIC (chloroplasts) pipeline is impaired.
Why don’t the ER or Golgi qualify as organelles that can grow and divide to reproduce themselves?
A common exam trap is to treat “having division machinery” as equivalent to “self-replicating organelles.” The nucleus, ER, Golgi, and lysosomes are typically inherited as existing structures during cytokinesis rather than assembled from scratch or autonomously expanded by binary fission.
If you removed all mitochondria from a cell, could the cell regenerate new mitochondria just from nuclear instructions?
When mitochondria are depleted, cells cannot rebuild them from scratch using only nuclear gene expression, because the process requires preexisting mitochondrial components to start organelle biogenesis. That irreversibility is why mitochondria and chloroplasts are considered truly autonomous in a way that peroxisomes are not.
How can you tell whether an organelle is truly reproducing versus just fragmenting?
You generally cannot count “division” events as proof of autonomous organelle replication. A mitochondrion might fragment due to altered fission control without a true increase in organelle mass or complete completion of biogenesis, so the functional test is whether new membrane/protein content and DNA replication are coordinated with fission.
Is chloroplast division strictly the same as mitochondrial fission, or does light change the requirements?
Chloroplast division depends on additional environmental and internal state inputs, especially light. Without appropriate light conditions, chloroplast DNA synthesis and timing cues can be disrupted, so you may see slower or uncoupled division cycles even if the core protein rings are present.
What are concrete examples of how the nucleus “holds the throttle” on organelle reproduction?
Another frequent confusion is thinking organelles are independent decision-makers. In mitochondria, nuclear control examples include TFAM levels, and in chloroplasts, retrograde signaling adjusts nuclear gene expression based on chloroplast redox and ROS status, so replication is gated by host regulation.
Can mitochondria or chloroplasts divide if a cell is energy-starved or missing membrane precursors?
If the cell lacks energy or membrane building blocks, organelle reproduction is throttled because biogenesis requires more lipids, more proteins, and enough ATP for translation and active transport. Even with intact division machinery, insufficient cellular resources will prevent the buildup needed for successful daughter organelles.
Why might DRP1-mediated pinching fail to produce new functional mitochondria even if DRP1 is present?
Mitochondrial division is tightly coupled to upstream growth steps, so if TFAM-driven mtDNA replication or protein import does not keep pace, newly forming constriction sites can fail to yield stable daughter mitochondria. In other words, division is not just a mechanical pinch, it is an end-stage event after growth and DNA replication have prepared the substrate.
Why do isolated mitochondria often fail to divide in vitro, even though they can divide inside cells?
In cell biology experiments, results depend on how organelles are prepared. Purified mitochondria lose the ability to divide quickly because they no longer receive the host-supplied proteins, membrane context, and energy support that a living cell provides, so in vitro observations can differ from in vivo autonomy.
Next Article
How Do Cells Grow and Reproduce to Maintain Homeostasis
Learn how cells grow and divide in controlled steps, balancing cell cycle, DNA replication, and signals to keep homeosta
