Yeast cells grow by building up mass and then reproducing through a process called budding, where a small daughter cell pinches off from the parent. In the right conditions (warm temperature, available glucose, near-neutral to slightly acidic pH), Saccharomyces cerevisiae can double its population roughly every 90 minutes. This process is what allows yeast to can grow and reproduce under favorable conditions. But yeast growth is not unlimited: ethanol accumulation, nutrient depletion, pH shifts, and osmotic stress all put a hard ceiling on how far a population can expand. Understanding what drives growth and what stops it gives you the practical control you need, whether you're running a lab experiment, making bread, or just trying to understand how single-celled organisms tick.
How Do Yeast Cells Grow and Reproduce: Step by Step

What 'growth' actually means for a yeast cell
It's worth splitting the word 'growth' into two separate ideas before going any further, because they're easy to conflate. The first is cell growth: an individual yeast cell getting physically larger, accumulating proteins, lipids, ribosomes, and other biomolecules. The second is population growth: the total number of cells in a culture increasing over time. Both matter, and they're connected, but they don't always move together.
A yeast cell that's metabolically active (churning out CO2 and ethanol from sugar) is not necessarily dividing. This distinction trips up a lot of beginners who see vigorous bubbling in a flask and assume the cell count must be skyrocketing. Fermentation is a metabolic activity; reproduction requires a full cell cycle. You can have one without the other ramping up proportionally, which is exactly why measuring actual cell number (or optical density calibrated against cell counts) matters if you want to track real population growth.
Budding: the main way yeast reproduce
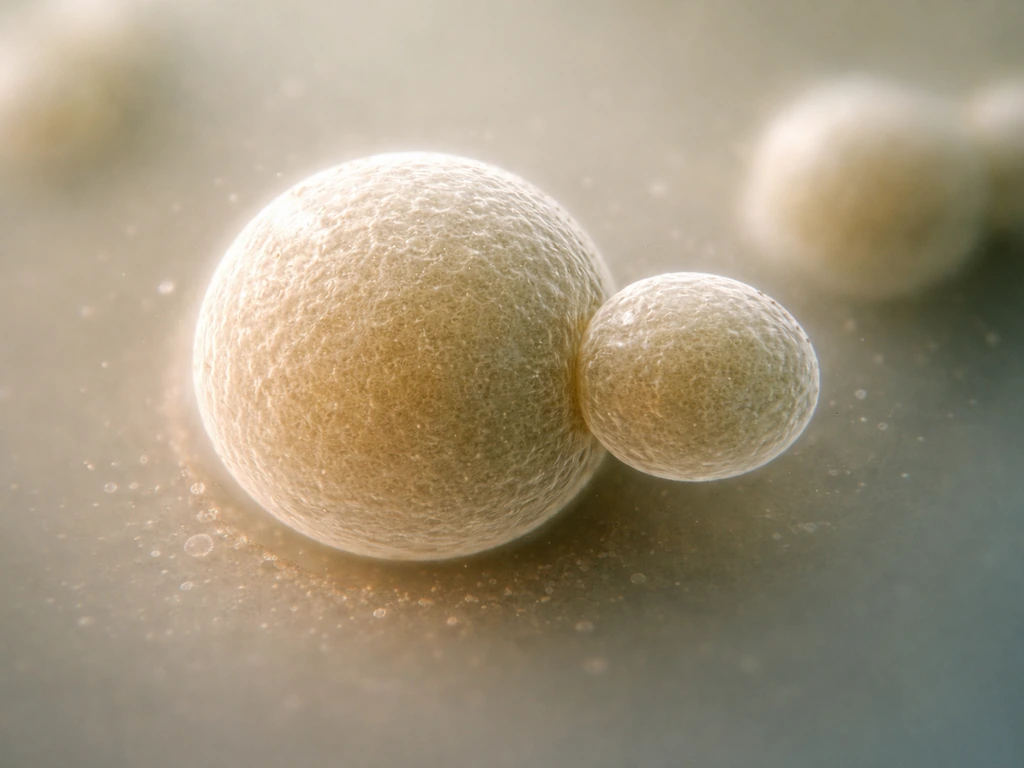
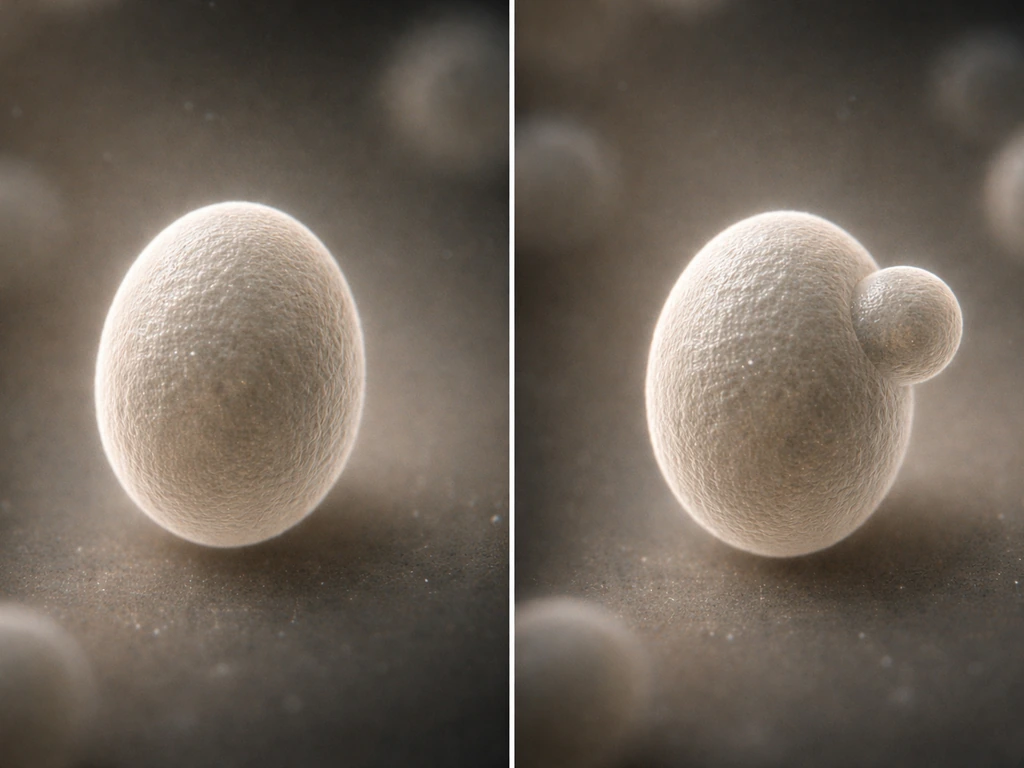
Saccharomyces cerevisiae reproduces primarily by budding, a form of asexual reproduction that's distinct from the symmetric binary fission you see in bacteria. The parent cell grows a small protrusion, or bud, at a specific site on its surface. The nucleus divides, one copy migrates into the bud, and then the bud separates. These organelles can grow and divide to reproduce themselves, which is why the cell's internal division machinery matters for population expansion. The parent and daughter are not equal in size: the parent keeps most of its volume and can bud again, while the daughter starts small and must grow before it can bud itself. Each division leaves a bud scar on the parent cell wall, and a parent can only accumulate a limited number of these before it ages out and stops dividing.
The decision to bud is controlled by the yeast cell cycle, and the critical checkpoint is called START. Think of START as a gate at the boundary between G1 (growth phase) and S phase (DNA replication). The gate is held shut by a protein called Whi5, which inhibits the transcription factors SBF and MBF. Those transcription factors need to be active to kick off the gene expression program that drives the cell into division. The lock on the gate is released when a cyclin called Cln3 partners with the main yeast CDK (a kinase called Cdc28) and phosphorylates Whi5, booting it out of the nucleus. In budding yeast, the main CDK Cdc28 partners with nine cyclins total, including three G1 cyclins (Cln1, 3), and these G1 cyclins regulate Start Cdc28 with nine cyclins total and Start regulation by G1 cyclins. Once Whi5 is gone, SBF and MBF activate the late G1 cyclins Cln1 and Cln2, and the cycle is committed.
The key practical implication: a yeast cell has to reach a critical size before it crosses START. Nutrients and energy supply drive that growth. When nutrients are plentiful, cells hit the size threshold faster and cycle more rapidly. When nutrients are scarce, the START transition can actually become reversible, meaning a cell that was heading toward division can be pushed back into a quiescent state. This is the cell's way of not wasting resources dividing when conditions don't support it.
The yeast cell cycle in brief
- G1 phase: the cell grows in size; nutrient signals determine whether and when START is crossed
- START (G1/S transition): Cln3-Cdc28 phosphorylates Whi5, releasing SBF/MBF and committing the cell to division
- S phase: DNA is replicated; bud emergence begins
- G2/M phase: the spindle pole body (yeast's equivalent of a centrosome) duplicates, and the mitotic spindle assembles
- Mitosis and cytokinesis: the nucleus divides, chromosomes segregate, and the bud pinches off as a new cell
How nutrients, energy, and oxygen shape yeast growth

Glucose is yeast's preferred carbon source, and the cell has elaborate molecular machinery to prioritize it. When glucose is abundant, a repressor protein called Mig1 (activated by the nutrient-sensing kinase Snf1) suppresses genes needed for alternative carbon sources and for respiratory metabolism. The cell basically ignores the mitochondria and runs almost entirely on fermentation, converting glucose to ethanol and CO2. This is the Crabtree effect, and it's why yeast bubbles enthusiastically in a sugary medium even when oxygen is available.
When glucose runs out, the Snf1 kinase releases Mig1's repression and the cell shifts to respiratory metabolism, burning the ethanol it produced during fermentation. This transition is called the diauxic shift, and it's visible as a second, slower growth phase after the initial rapid growth. Growth rate drops significantly after the shift because respiration, while more energy-efficient per glucose molecule, is slower in terms of biomass production per unit time than all-out fermentation.
Oxygen availability matters here. With oxygen, yeast can run aerobic respiration and handle the shift after glucose depletion. Without oxygen (strictly anaerobic conditions), yeast is locked into fermentation and will stop growing once glucose is gone and toxic ethanol builds up. For maximum cell number and sustained population growth, aerobic conditions with sufficient but not excessive glucose are ideal. If you just want fast fermentation (for bread rising or brewing), anaerobic plus high glucose works fine, but it's not the same as maximizing cell reproduction.
The conditions that control how fast yeast reproduce
Four environmental variables do most of the work in controlling yeast reproduction rate: temperature, pH, osmotic conditions, and the presence of inhibitory compounds. Getting these right is the difference between a sluggish, stalled culture and a population doubling every 90 minutes.
Temperature
S. cerevisiae grows optimally around 30 to 35 °C in most lab strains, with a doubling time near 90 minutes at 30 °C in rich media. Below about 15 °C, growth slows dramatically as enzyme kinetics drop off. Above 37 °C, heat stress responses kick in and growth slows; above 40 to 42 °C, most strains stop growing entirely. For a practical home or lab setup, maintaining 28 to 32 °C will give you the fastest reliable growth without heat stress complications.
pH
Yeast prefers a mildly acidic environment. Growth rate is relatively stable across pH 4.5 to 6.0, making this the comfortable window to work in. As pH rises toward 7.0 and above, growth becomes less efficient. Under salt stress conditions, the acceptable pH range narrows further, to roughly pH 4.0 to 5.0. If your culture is stuck, check pH first: fermentation itself produces organic acids and CO2 that can drag pH down over time, which may actually self-limit growth in unbuffered media.
Osmotic stress
High concentrations of sugars or salts pull water out of yeast cells through osmosis, activating a stress response pathway called the HOG (High Osmolarity Glycerol) pathway. The cell diverts resources to producing glycerol as an osmolyte to counteract the external water potential, and this directly reduces the single-cell growth rate and caps maximum cell size. Practically, this means very high sugar concentrations (as in honey or concentrated wort) slow yeast growth considerably even if all other conditions look fine.
Ethanol and other inhibitors

Ethanol is yeast's own metabolic waste product, and it's toxic to the cells producing it. Ethanol concentrations of around 8 to 9% (v/v) begin to clearly distinguish tolerant from non-tolerant strains in growth assays. At concentrations in the 7 to 10% range, ethanol disrupts membrane integrity and intracellular pH homeostasis in non-adapted strains. Beyond about 12 to 15% (v/v), even tolerant strains cease reproducing. This is why wine fermentations stall in that alcohol range without any external intervention.
Why yeast growth always hits a wall
If you plot yeast cell number over time in a closed system (a flask with no fresh nutrients added), you'll see four phases: a lag phase while cells adjust to their environment, a log (exponential) phase where cell number doubles repeatedly, a stationary phase where division rate equals death rate and the count plateaus, and eventually a death phase. The carrying capacity of that system, the maximum cell number it can support, is determined by all the constraints above acting together.
In stationary phase, it's not that cells stop all activity. It's that resources are too depleted and inhibitors (especially ethanol and organic acids) are too concentrated for net growth to occur. The cell count looks stable on a plate count or OD reading, but underneath that stability is a continuous process of some cells dying and a matching fraction dividing. Chronological aging in yeast is defined precisely by this loss of viability in non-dividing stationary-phase cells: the longer cells sit in depleted medium, the more their viability drops even though the apparent count hasn't changed much.
The conceptual point here matters: no closed system allows unlimited growth. The same carrying-capacity logic applies to yeast in a flask, bacteria in a petri dish, and human populations in a resource-limited environment. Growth rates are not intrinsic constants; they're negotiated in real time between the organism's biology and the environment's supply of what that organism needs.
How to set up and verify yeast growth yourself
Whether you're doing a classroom experiment or a home kitchen test, the core setup is the same: give yeast a defined medium, control the key variables, and measure over time. Here's a practical walkthrough.
Basic setup

- Dissolve 5 to 10 g of sugar per 100 mL of water; add a pinch of yeast nutrients (or use standard YPD broth if you have lab access)
- Adjust or verify pH: aim for 4.5 to 5.5 using a pH strip or meter
- Inoculate with a small amount of active dry yeast or a liquid culture at low density (OD600 around 0.05 to 0.1 if you have a spectrophotometer)
- Incubate at 28 to 32 °C with gentle agitation if possible (shaking improves oxygen transfer and gives faster aerobic growth)
- Measure every 30 to 60 minutes during the expected exponential phase
Measuring growth: what to actually track
Optical density at 600 nm (OD600) is the standard quick measurement in lab settings. It measures how much light is scattered by cells in suspension, which correlates with cell density. As a rough calibration, OD600 = 1 corresponds to roughly 3 x 10^7 cells per mL in S. cerevisiae, though this number varies with strain and instrument setup and should be validated with a plate count if accuracy matters. OD600-based turbidity is commonly used to determine growth phase and cell number in actively growing cultures, but the OD-to-cell calibration should be empirically validated (for example, by plate counts). OD600 doesn't tell you whether cells are alive, only that particles are scattering light, so a culture in which cells are dying and the dead cells haven't lysed yet can still read a high OD.
For viability, the simplest accessible test is a methylene blue stain: living cells reduce the dye and stay colorless, while dead cells stain blue. In a lab context, propidium iodide (PI) staining is more accurate because PI only penetrates cells with compromised membranes, letting you clearly count dead versus living cells under a fluorescence microscope or with a flow cytometer. Fluorescein diacetate (FDA) is another option: it's converted to a fluorescent product only inside metabolically active cells with intact membranes.
Troubleshooting: why growth might be slow or stalled

| Symptom | Likely cause | What to adjust |
|---|---|---|
| No growth after 2–3 hours | Dead or inactive starter culture; temperature too low | Use fresh yeast; confirm incubation temperature is 28–32 °C |
| Slow growth, lots of bubbling | Fermentation active but cell division limited (anaerobic, high sugar) | Add oxygen (swirl or aerate); dilute sugar if above 15% |
| Growth stops mid-log phase | Glucose depleted or ethanol inhibition | Add fresh glucose or dilute the culture to lower ethanol concentration |
| Culture crashes after initial growth | pH dropped too far or nutrient exhaustion | Buffer the medium; add fresh nutrients or dilute into fresh medium |
| OD rises but viability test shows mostly dead cells | Cells dying faster than dividing (late stationary or death phase) | Subculture into fresh medium immediately; reduce initial cell density |
| Growth slower than expected at all phases | Temperature off; osmotic stress from high sugar/salt | Check thermometer calibration; reduce solute concentration |
Quick checklist for robust yeast reproduction
- Temperature: 28 to 32 °C for optimal doubling time near 90 minutes
- pH: 4.5 to 5.5; monitor and buffer in longer experiments
- Glucose: present but not excessive (1 to 10% is a practical range for most lab cultures); very high concentrations create osmotic stress
- Oxygen: aerobic conditions support sustained population growth; anaerobic favors fermentation over cell division
- Ethanol: keep below 8% (v/v) in the culture for active cell division, especially with non-tolerant strains
- Nitrogen and micronutrients: often overlooked; pure sugar water gives poor results; add yeast extract or nitrogen source for real growth
- Starting cell density: too high means you enter stationary phase quickly; start dilute and let the exponential phase run
Yeast as a model for how all growth hits limits
Yeast isn't just a convenient lab organism. It's one of the clearest windows into the universal rules that govern growth across all of biology. The same logic that applies to a flask of yeast applies to how human cells grow and reproduce, what stem cells need to divide, and why no living system expands indefinitely without external input. Before human cells can grow and reproduce they need sufficient resources and conditions to pass key growth and division checkpoints what stem cells need to divide.
Every cell, from yeast to human, ties cell division to a checkpoint that requires sufficient size and energy. In the same way, stem cells must reach the right size and have enough nutrients, growth factors, and oxygen to pass checkpoints and keep dividing sufficient size and energy. In human cells, growth and reproduction likewise depend on having enough nutrients, energy, and growth signals to pass cell-cycle checkpoints what do human cells need to grow and reproduce. In yeast, that's the Whi5/SBF/Cln3 system at START. In human cells, an analogous system (the retinoblastoma protein, Rb, functioning much like Whi5) controls the G1/S transition. The specific molecular players differ, but the logic is the same: grow to a threshold, confirm resources are available, then commit to dividing. If resources fail, pause or abort.
And in every system, waste accumulation is a hard constraint. Yeast hits its wall because of ethanol. A tissue hits its wall because oxygen and glucose diffusion limits how deep nutrients can reach. A population hits its wall because food, space, or some other resource becomes limiting. The carrying capacity of a system isn't a vague ecological concept; it's the direct physical and chemical consequence of the same constraints you can measure in a yeast flask on your bench today.
If you want to explore these ideas further, the same principles show up when you look at what human cells need to grow and reproduce, or at how cells more broadly grow and reproduce to maintain homeostasis in a tissue. Yeast is a great starting point because the variables are easy to manipulate and the feedback is fast enough to see in a single afternoon experiment. Start a culture this morning, check it every hour, and by the time the exponential phase peaks and stalls, you'll have watched the full arc of growth, saturation, and limit in real time.
FAQ
How can I tell whether yeast is actually reproducing or just fermenting?
Track viable cell number or calibrated OD rather than CO2 bubbling. Fermentation can be high while division is paused or slowed if cells have not met the size threshold for START, if ethanol is already stressing them, or if nutrients are depleted. A quick cross-check is OD600 plus a viability stain (methylene blue or PI) to see whether cell death is offsetting any apparent growth.
Why does my culture stall before glucose is completely gone?
Common causes are pH drift (especially in unbuffered media), osmotic stress from high sugar or salt, oxygen limitation even under “aerobic” conditions, and early ethanol inhibition in high-glucose runs. Check pH and measure or estimate ethanol if possible, then reduce sugar concentration or improve aeration and mixing (more headspace or better shaking) before rerunning.
Does oxygen always increase yeast growth rate?
Oxygen helps during the diauxic shift and supports respiration after glucose depletion, but oxygenation alone will not guarantee faster division if glucose is in excess or if ethanol and pH have already pushed cells into stress. Also, “aerobic” cultures require adequate oxygen transfer, which depends on shaking speed, flask volume, and surface area, not just the presence of air.
What sugar concentration gives the best maximum cell number, not just fast fermentation?
There is a tradeoff. Very high glucose can raise osmotic stress and increase ethanol accumulation, both of which reduce division rate and cap carrying capacity. For maximizing cell number, aim for a moderate glucose level, keep temperature controlled, and ensure conditions allow the diauxic shift rather than staying stuck in fast-fermentation stress.
Why is the lag phase longer at the start sometimes?
Lag increases when cells are transferred from a different medium, when starting cells are stressed (e.g., high ethanol or low nutrients), or when temperature and pH are not matched to their prior environment. Let cells adapt to the new conditions, and consider pre-culturing into the same medium for more consistent START timing.
Is OD600 a reliable way to calculate doubling time?
It can be used, but you must use a calibration curve for your strain, instrument, and growth medium, and you should stay within the linear OD range of your spectrophotometer. OD600 also does not distinguish live from dead cells, so doubling time estimates can be misleading if viability drops while light scattering stays high.
How do I minimize measurement error with viability stains?
Perform staining quickly after sampling because prolonged holding can change cell membrane integrity and dye behavior. Use proper controls (untreated live and heat-killed dead cells) to set thresholds, and mix well so you do not undercount cells that clump.
What causes daughter cells to be able to bud repeatedly, while some cells stop budding?
Budding capacity depends on reaching a committed cell-cycle state and sustaining internal division machinery. If a cell accumulates too many bud scars, if resources are depleted, or if stressors like ethanol and osmotic pressure damage homeostasis, the START pathway may not be completed, leading to non-dividing cells even if they remain metabolically active.
How can I tell whether the problem is temperature versus pH versus osmotic stress?
Change one variable at a time and include a matched control. Temperature shifts rapidly affect enzyme kinetics, pH drift can creep over time due to fermentation byproducts, and osmotic stress scales with solute concentration. If growth is slow immediately after setup, suspect temperature or starting-state mismatch; if it slows after hours, pH and ethanol accumulation are prime suspects.
Why does growth slow after the diauxic shift, even though respiration is more efficient?
Respiration yields more energy per glucose molecule, but biomass production per unit time can still be slower because the metabolic program changes, the cell must retool gene expression, and growth becomes limited by how quickly the new pathway can ramp up. That is why the second phase is often slower despite improved efficiency.
What happens if I keep the culture in a closed flask but keep adding sugar?
Adding fresh sugar changes the environment, effectively increasing carrying capacity and delaying stationary phase, but it can also keep ethanol production high if the culture stays in fermentation. To truly improve sustained reproduction, also consider oxygen transfer and buffering, because waste accumulation (ethanol, organic acids) often becomes the main limiter.
Can Grow and Reproduce: Conditions, Limits, and Proof
Learn what it means to can grow and reproduce, conditions needed, why limits stop it, and how to verify proof.
