Stem cells grow in two places: inside the body, tucked into specialized microenvironments called niches, and in the lab, where scientists recreate just enough of those conditions to keep them dividing without losing their identity. Neither location is passive. Whether it's a pocket of bone marrow or a coated plastic flask, stem cells only keep dividing when the chemistry around them says it's okay to do so. Get those signals wrong and the cells either stop growing, die, or turn into something else entirely.
Where Do Stem Cells Grow: Body Niches and Lab Culture

Marcus Whitmore
25 Apr 2026
What we mean by "stem cells" and "grow"
Not all stem cells are the same, and that matters a lot when you're asking where they grow. The NIH groups them into two broad buckets: pluripotent stem cells (embryonic stem cells and induced pluripotent stem cells, or iPSCs) and somatic or adult stem cells. NIH/NIGMS explains that somatic (adult) stem cells are multipotent and that induced pluripotent stem cells are created in the laboratory, alongside embryonic stem cells as a separate category of pluripotent stem cells somatic or adult stem cells. Pluripotent cells can, in principle, become almost any cell type in the body. Adult stem cells are more specialized, differentiating mainly into the tissue types of the organ where they live.
"Growing" is also a term worth unpacking. For stem cells, growth isn't just getting bigger. It means self-renewal: the ability to divide and produce daughter cells that stay stem cells, keeping the pool replenished. At the same time, some daughters take a different path and differentiate, committing to become a specific cell type like a blood cell, a gut lining cell, or a hair follicle cell. The balance between those two outcomes, self-renewal versus differentiation, is the central drama of stem cell biology.
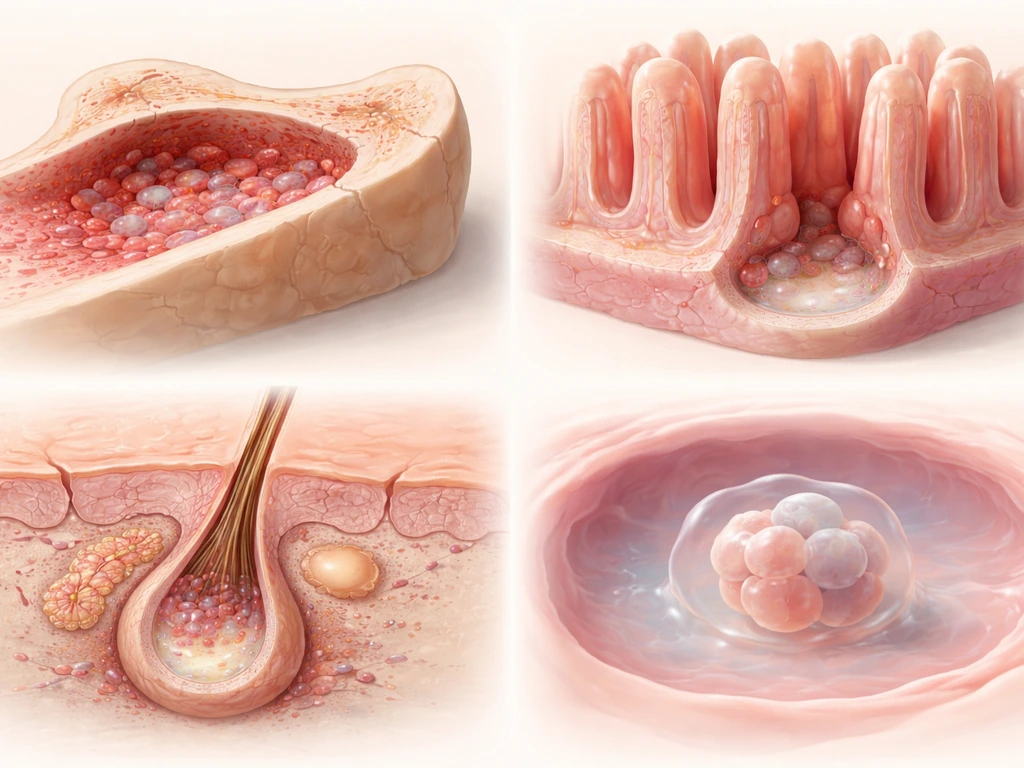
Where stem cells live in the body: the niche concept

Think of a stem cell niche the way you'd think of a very specific garden bed. Pull a seedling out and put it on bare concrete and it won't thrive, even though it's the same plant. Stem cells are the same: they depend on signals, physical contacts, and local chemistry that only exist in that particular spot.
Bone marrow: the hematopoietic niche
Hematopoietic stem cells (HSCs), the ones that produce every type of blood cell, sit in the bone marrow. About 80% of them park themselves right next to sinusoidal blood vessels, the slow-flowing capillary-like channels inside marrow. The rest cluster near arterioles or in the endosteal region, the zone just inside the bone surface. The niche isn't just a parking spot; it actively talks to the stem cells. Osteoblasts lining the bone surface and reticular cells throughout the marrow secrete signals like CXCL12 and Stem Cell Factor (SCF), which regulate homing, retention, and the decision to stay quiescent or start dividing.
The gut: intestinal crypts
The lining of your small intestine is completely replaced roughly every five days, one of the fastest renewal rates in the body. That regeneration is driven by Lgr5-positive stem cells sitting at the base of tube-like structures called crypts. Their immediate neighbors are Paneth cells, which act as the niche support crew. Paneth cells supply EGF, Wnt3, and Notch ligands that keep Lgr5+ stem cells dividing and undifferentiated. Multiple signaling pathways, including Wnt, BMP, Notch, Hedgehog, and EGF, coordinate to control whether crypt stem cells self-renew or head up toward the villus tip and differentiate.
Hair follicles: the bulge
Hair follicle stem cells occupy a region called the bulge, a physical swelling on the follicle roughly two-thirds of the way down from the skin surface. These cells are marked by proteins like K15, CD34, and integrin alpha-6, along with transcription factors including Sox9, Lhx2, and Nfatc1. They sit in relative quiet between hair cycles and activate during the growth phase, demonstrating how niche control can be temporally as well as spatially precise.
Early embryo: where embryonic stem cells come from
Embryonic stem cells (ESCs) are isolated from the inner cell mass of a blastocyst, a roughly 5-to-7-day-old embryo. This is a transient structure in development, not a permanent niche like bone marrow. Once ESCs are removed and placed in culture, there is no "natural" place they grow in the adult body. Induced pluripotent stem cells (iPSCs) skip that origin entirely: they're made by reprogramming ordinary adult cells (like skin fibroblasts) using transcription factors such as OCT4, NANOG, KLF4, and c-Myc, essentially rewinding the clock on cellular identity.
Growing stem cells in the lab: what culture actually looks like

If the niche is a garden bed, then a cell culture flask is a hydroponic setup: you strip away the soil and try to pipe in everything the plant needs artificially. It works, but you have to get the recipe right and keep it consistent.
Feeder cells vs. feeder-free systems
Early embryonic stem cell culture relied on growing cells on top of a layer of mouse embryonic fibroblasts (MEF feeder cells) that had been treated so they couldn't divide. The feeders secreted a mix of growth factors that mimicked niche support. For mouse ESCs, simply adding LIF (Leukemia Inhibitory Factor) at around 1,000 U/mL to serum-containing DMEM on a gelatin-coated surface achieves similar results without feeders.
For human pluripotent stem cells today, the standard has largely shifted to fully defined, feeder-free media like Essential 8 (E8), used on surfaces coated with vitronectin or Matrigel. You can also design lab systems where stem cells grow on plastic surfaces if you provide the right coatings, media, and signals. E8 is also xeno-free, meaning it contains no animal-derived components, which matters for clinical manufacturing.
Oxygen: the overlooked variable

Most labs default to atmospheric oxygen, about 21%, but stem cell niches in the body are hypoxic. Reducing O2 tension to somewhere in the 2-5% range reduces spontaneous differentiation in human ESC cultures and activates HIF (hypoxia-inducible factor) signaling that supports the undifferentiated state. If you're troubleshooting a culture that keeps differentiating unexpectedly, oxygen level is worth checking.
Expanding blood-forming stem cells ex vivo
HSCs are famously hard to expand outside the body without triggering differentiation. Serum-free media supplemented with a cytokine cocktail, typically including SCF, TPO, FLT3 ligand, and sometimes IL-3, IL-6, or Notch ligand Delta-like 1, can maintain hematopoietic progenitor activity. Getting the cytokine combination right is the subject of ongoing research, and what works for one application doesn't automatically work for another.
What controls whether a stem cell divides or differentiates
The decision between self-renewal and differentiation isn't random. It comes down to the balance of signals a stem cell receives at any given moment. In the gut, Wnt signaling from Paneth cells keeps Lgr5+ cells in a stem state; move away from those cells and BMP signaling ramps up, pushing differentiation. For pluripotent cells in culture, pluripotency markers like OCT4, NANOG, and SOX2 are actively maintained by growth factors in the media. Once those signals drop, or once differentiation cues arrive, the transcriptional program shifts. Practically, this is why you need daily medium changes in E8-based iPSC culture: the growth factors degrade and cells start drifting if you skip a day.
Notch signaling deserves a special mention because it does double duty. In the intestinal crypt, Notch positive feedback is essential for stem cell self-renewal. In hematopoiesis, Notch ligand Delta-like 1 is part of the ex vivo expansion toolkit. The same pathway plays different roles depending on context, which is a recurring theme in stem cell biology and a reason why copying protocols from one cell type to another often doesn't work straight out of the box.
Why stem cells can't just grow forever: the real limits
This connects directly to a bigger principle: biological growth always has constraints. Stem cells are no exception. In the body, niche size is finite. The bone marrow can only physically accommodate so many HSCs, and the crypt base holds only a defined number of Lgr5+ cells. Remove the niche signals and cells either differentiate, become quiescent, or die.
In culture, the constraints are different but equally real. Monolayer culture is limited by surface area: once cells hit 100% confluency, you have to split the culture or differentiation climbs. A common guidance threshold is keeping differentiation below roughly 20%, and passaging on schedule rather than letting plates overgrow is one of the main ways to control that. Beyond physical space, longer-term expansion hits a biological wall.
Telomeres shorten with each division, and once they get short enough, stem cells enter senescence, reducing division capacity and eventually triggering apoptosis. In mesenchymal stem cells cultured long-term, telomere erosion correlates with oxidative stress and genomic instability, including aneuploidy. That's a genuine upper limit on how many times you can passage a cell line before quality degrades.
Reprogramming to iPSCs actually resets telomere length somewhat, which is part of why iPSCs can be expanded for longer than many primary adult stem cell cultures. But that isn't unlimited either: redifferentiation and extended passaging still cause telomere shortening over time.
Choosing the right stem-cell source: adult, embryonic, or iPSC
If you're trying to figure out which type of stem cell is relevant to your question or project, here's a practical comparison.
| Source | Where they grow in the body | Pluripotency | Practical culture difficulty | Key clinical concerns |
|---|---|---|---|---|
| Adult stem cells (e.g., HSCs, intestinal, hair follicle) | Tissue-specific niches (marrow, crypts, bulge) | Limited (tissue-restricted) | Moderate to high; hard to expand without differentiation | Low immunogenicity if autologous; limited differentiation range |
| Embryonic stem cells (ESCs) | Inner cell mass of blastocyst (transient); no adult niche | Full pluripotency | Moderate with feeder-free/E8 systems | Ethical issues; immune rejection in allogeneic use; teratoma risk |
| Induced pluripotent stem cells (iPSCs) | No natural body location; made in the lab | Full pluripotency | Moderate with defined media; requires careful QC | Immunogenicity possible; genomic instability risk; teratoma risk if undifferentiated cells remain |
For most research questions about a specific tissue (bone, cartilage, blood), adult stem cells from that tissue are the most relevant starting point. For questions about growing virtually any cell type, pluripotent cells (especially iPSCs, since they sidestep embryo ethics) are the more versatile tool. iPSCs also let you work with patient-derived cells, which matters enormously for disease modeling.
One safety issue worth knowing about if you're reading clinical claims: residual undifferentiated pluripotent cells can form teratomas after transplantation. Teratoma formation in immune-deficient mice is actually the gold-standard assay for confirming pluripotency in a cell line, so the same property that makes these cells valuable also creates a safety hurdle in therapy.
However, ISSCR standards note that xenograft (teratoma) assays are not required when equivalent information can be derived from in vitro pluripotency assays Teratoma formation in immune-deficient mice is actually the gold-standard assay for confirming pluripotency in a cell line. Responsible lab workflows include checking pluripotency markers like SSEA-4 and TRA-1-60 by flow cytometry, running PluriTest (a bioinformatics assay using genome-wide expression data), verifying karyotype stability, and following ISSCR guidelines on genomic characterization.
Practical next steps if you're solving this problem today
Here's how to orient yourself quickly depending on your goal.
- Identify your cell type first. Are you asking about blood stem cells, gut stem cells, neural stem cells, or pluripotent stem cells? The niche, the culture conditions, and the relevant signals are completely different for each.
- Learn the vocabulary of the field before evaluating any protocol or product. The key terms to look for are: niche, self-renewal, differentiation, feeder-free, defined media, passage number, and pluripotency markers. If a protocol doesn't specify these, it's incomplete.
- For pluripotent cells in culture, start with Essential 8 on vitronectin-coated surfaces, use ROCK inhibitor when passaging to improve survival, change media daily, and keep O2 at 5% if your incubator supports it.
- For HSC or blood progenitor expansion, look for serum-free protocols using SCF, TPO, and FLT3 ligand as a base cytokine combination, then adjust based on your specific application.
- Validate your cells regularly. Surface markers (SSEA-4, TRA-1-60 for pluripotent cells), morphology, karyotype checks, and expression of pluripotency transcription factors are not optional QC steps, they're how you know your stem cells are actually still stem cells.
- Check passage number on anything you're expanding. Telomere shortening and genomic drift are real over long-term culture. If you're above passage 30-40 on a human iPSC line without recent karyotyping, treat the data from those cells with caution.
If you're interested in what stem cells can actually build once they do differentiate, questions like whether they can generate cartilage, bone, or other structures connect directly to how the differentiation cues work and what physical scaffold they need to organize properly. In some settings, stem cells can stem cells grow bone by differentiating into bone-forming lineages when the right signals and scaffold are provided.
In practice, whether you can grow stem cells depends on choosing the right cell type and recreating the signals and conditions that keep them undifferentiated can you grow stem cells. This is one reason researchers spend so much effort designing the right differentiation cues and scaffolds to guide stem cells toward cartilage generate cartilage. Those are natural extensions of understanding where stem cells grow and what controls the switch between self-renewal and committed cell identity.
FAQ
Do stem cells “grow” in just one place, or can they live in multiple niches at once?
They can depend on the cell type. For example, hematopoietic stem cells are distributed among several marrow regions (near vessels and near the bone surface). That means a single stem cell population can be supported by multiple microenvironments, and moving conditions (or physically relocating cells in the body) can shift behavior toward quiescence or differentiation.
If ESCs have no adult body niche, why do pluripotent cells still stay undifferentiated in the lab?
In culture, pluripotency is maintained by continuously supplying the signaling environment the niche would provide during development (growth factors plus controlled surface and oxygen). Without that ongoing “niche-like” input, pluripotent cells rapidly drift into differentiation even if they are still alive.
Why do some stem cell cultures differentiate even when the media recipe seems correct?
Oxygen level and cell density are common hidden variables. Human pluripotent cultures often require hypoxic conditions (roughly 2 to 5% O2) to reduce spontaneous differentiation, and cells also respond to how crowded they are, because neighbors change local signaling and nutrient use.
How do I know whether a stem cell line is actually losing “stemness,” not just slowing down?
Look for identity markers and functional readouts together. A culture may appear viable but shift transcriptionally, changing pluripotency or progenitor programs. For pluripotent cells, combining marker checks (for example, SSEA-4 and TRA-1-60 by flow) with genome stability (karyotype, PluriTest-style expression profiling) helps distinguish true maintenance from early drift.
What happens to adult stem cells when the niche signals are disrupted?
Many adult stem cells rely on retention and quiescence cues. If those signals drop, they are more likely to exit quiescence, differentiate prematurely, or undergo stress. In expansion work, this is why ex vivo HSC growth is sensitive to the exact cytokine balance and why the same approach does not universally transfer to other adult stem cell types.
Why is passaging schedule so important for stem cells in culture?
Overgrowing a monolayer changes the microenvironment around cells. Confluency affects nutrient gradients, waste buildup, and cell-cell signaling, which can push differentiation upward. Practical practice is to passage before cultures reach a point where differentiation begins to climb and then keep the schedule consistent to avoid “signal drift” over time.
Can you expand hematopoietic stem cells (HSCs) indefinitely in culture?
No, continuous expansion is limited by biology. Even if cytokines support progenitor activity, long-term cultures face a ceiling from telomere shortening, oxidative stress, and genomic instability. Reprogramming to iPSCs can extend proliferative potential relative to many primary adult stem cell setups, but iPSCs still eventually accumulate limitations with extended passaging.
Is it safe to transplant pluripotent stem cell-derived products if not all cells are fully differentiated?
Residual undifferentiated pluripotent cells are a key safety risk because they can form teratomas after transplantation. That is why responsible workflows include pluripotency marker assessment and genomic characterization before clinical use, not just relying on differentiation time.
Why doesn’t a differentiation protocol for one stem cell type work for another?
Because the same pathways can play different roles depending on context. Notch is a good example, it supports intestinal crypt self-renewal and also contributes to hematopoietic expansion when paired with the right ligands. Transplanting a protocol without adapting the cell-specific biology often fails.
What is the biggest “first decision” when choosing which stem cells to use for a project?
Start by matching the cell state to your question. If you need a specific tissue lineage (bone, cartilage, blood), adult or tissue-relevant stem/progenitor sources are often the most direct. If you need broad developmental versatility or patient-specific modeling, pluripotent cells like iPSCs are usually the better starting point.
Next Article
Can You Grow Stem Cells? What It Means and How It Works
Learn what it means to grow stem cells, how culture conditions work, and why it requires specialized, regulated labs.
