Mitosis helps an organism grow in size primarily by increasing the total number of cells, not by making individual cells bigger. Each time a cell completes mitosis, it splits into two genetically identical daughter cells. Do that billions of times across a developing embryo or a healing wound, and you get larger tissues, bigger organs, and a physically larger organism. The size increase comes from accumulating more cells to build with, not from inflating the cells you already have.
How Mitosis Helps an Organism Grow in Size

Marcus Whitmore
28 Apr 2026
The core link between mitosis and organism growth
Think of building a wall. You can try to make each brick bigger, or you can just add more bricks. Organisms mostly go with more bricks. Mitosis is the process that produces those additional bricks: it takes one somatic cell and reliably copies it into two diploid daughter cells, each carrying the same genetic blueprint as the original. As this happens repeatedly across tissues, the total cell count rises, and rising cell count is what drives most of the increase in physical size you observe during development, childhood growth, and tissue repair.
There is an important distinction worth keeping clear. Cell proliferation means an increase in cell number over time. Cell growth means an increase in cell size or mass. Both can contribute to a larger organism, but it is proliferation through mitosis that does most of the heavy lifting in building tissues and organs from scratch. Mitosis is the biological engine for proliferation.
How mitosis increases cell number, and what that actually means for size
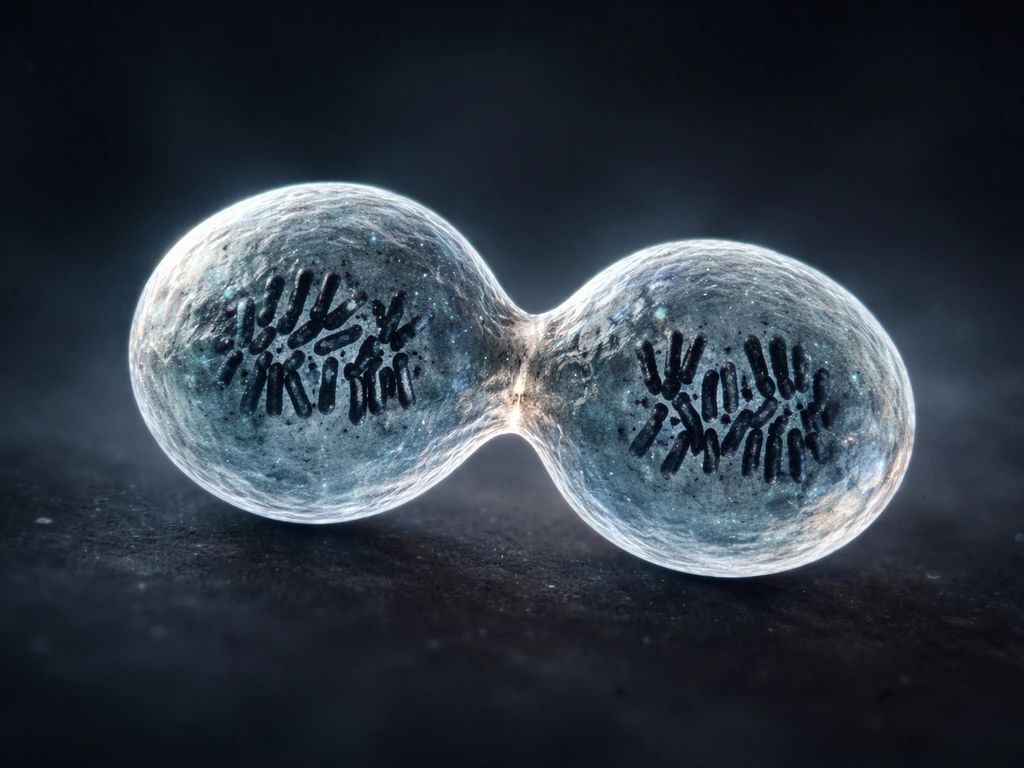
Mitosis is the M phase of the cell cycle. During this phase, a single parent cell divides its duplicated chromosomes equally and then physically splits, producing two daughter cells. Each daughter is genetically identical to the parent. This is not just copying information: it is physically doubling the number of membrane-bound units available to form tissue. Do this across millions of cells simultaneously during embryonic development, and a small cluster of cells becomes a recognizable organ within weeks.
Here is something slightly counterintuitive. The moment a cell divides, each daughter cell is actually smaller than the parent was just before division. Division by itself reduces individual cell size. What compensates is what happens between divisions: during interphase, cells grow back to a functional working size before they divide again. So organism-level size increase is really a product of the combination: cells grow a bit, divide, and the net result over many cycles is more cells total, building larger and larger structures.
Cell growth vs. cell division: which one drives what
This confusion trips up a lot of students, so it is worth spelling out clearly. Cell growth (increasing cell size or mass) and cell division (increasing cell number) are regulated independently. A cell can grow in mass without dividing, and cells can divide without proportionally increasing their mass. Research has shown that these two processes can be uncoupled at the molecular level, meaning 'divide more' does not automatically equal 'get bigger cells.'
| Process | What changes | Effect on organism size |
|---|---|---|
| Cell growth | Individual cell size/mass increases | Modest size increase; cells get larger but count stays the same |
| Cell division (mitosis) | Cell number increases | Major size increase; more cells available to build tissues |
| Both together | Cell count rises and cells maintain working size | Normal, sustained organismal growth during development |
During development, both processes are coordinated. A tissue that only divided without cells growing between divisions would produce smaller and smaller cells with each cycle until they became non-functional. A tissue where cells only grew but never divided would end up with a small number of giant cells that hit physical and metabolic limits fast. Normal growth requires the partnership: mitosis adds numbers, interphase growth maintains cell size and function.
The cell cycle: how cells decide when to divide

Mitosis does not happen constantly. Cells follow a structured cycle with checkpoints that work like permission gates. The canonical sequence is G1, S, G2, and then M phase. During G1, the cell grows and assesses whether conditions are right to commit to division. During S phase, DNA is replicated. G2 is a second growth and quality-check phase. Only after clearing all three checkpoints does the cell enter mitosis.
The molecular machinery behind those checkpoints centers on cyclin-dependent kinase complexes, or cyclin-CDK pairs. Specific cyclins rise and fall at defined points in the cycle. For example, cyclin E levels rise during late G1 and drop during S phase. Without the right concentration of activated cyclin-CDK complexes, the cycle cannot advance through a checkpoint. This is not a passive default: it is an active permission system.
Growth factors from outside the cell feed into this system directly. Cyclin D synthesis is triggered by growth factor signaling through the Ras-Raf-ERK pathway, and cyclin D persists as long as those signals are present. Remove the growth factors and cyclin D drops, stalling cells in G1. This is how the body connects tissue-level needs (is growth required here?) to cell-level behavior (should this particular cell divide now?). It is a remarkably elegant feedback loop.
There are also damage checkpoints. At the G1 checkpoint, cells check for DNA damage, sufficient nutrient reserves, and adequate size before committing to replication. At the G2/M boundary, the balance between Wee1 (which keeps Cdk1 inactive) and Cdc25 (which activates it) determines whether the cell enters mitosis or waits for DNA damage to be repaired. If damage is detected, p53 can trigger either a pause or programmed cell death. This is the cell cycle being protective, not permissive.
How tissues scale from cell numbers to organ size
An individual mitotic event is microscopic. So how does it translate to a liver, a leaf, or a leg? The answer is coordination at scale. Organ morphogenesis, the process of building organ shape and size during development, involves two coupled processes: increasing cell number through division and coordinating tissue shape and size through growth programs. Neither alone is enough. Organisms with many cells grow because cell proliferation via mitosis increases cell number and helps tissues build the larger structures they need organismal growth. Both processes are spatially and temporally coordinated so that the right cells divide at the right times in the right locations.
Stem cells play a central role in this scaling. In many tissues, a small population of stem cells divides to produce transit-amplifying cells, which then go through several rapid rounds of mitosis before differentiating into specialized cell types that stop dividing. The skin is a good example: stem cells in the basal layer divide, their progeny divide a few more times, and then those cells differentiate and migrate toward the surface, eventually being shed. This layered division strategy lets tissue maintain a large total cell count without requiring every cell to keep cycling.
Tissue homeostasis in adults depends on balancing cell addition through mitosis with cell loss through differentiation and death. Proliferation in development is carefully balanced with programmed cell death so that organs reach appropriate sizes and stop there. Some fully differentiated cell types, like mature neurons, are no longer capable of division at all. The 'growth phase' is built in to development and then deliberately shut off.
Why mitosis can't create unlimited size
If mitosis can keep adding cells, why do organisms stop growing? There are several converging constraints, and they operate at every scale from individual cells to entire organisms.
The surface-area-to-volume problem

Individual cells face a hard physical limit tied to diffusion. For a spherical cell, the surface-area-to-volume ratio is 3 divided by the radius. As a cell gets larger, that ratio shrinks, meaning each unit of cell volume has less membrane surface through which to exchange nutrients and waste. At some point, the cell interior becomes too large for diffusion to supply oxygen and remove carbon dioxide efficiently. This is one core reason cells divide rather than growing indefinitely: division resets the surface-to-volume ratio to a workable level for each daughter cell.
Nutrient and oxygen supply at the tissue level
Scaling up from individual cells to tissues, nutrient and oxygen supply become limiting factors for larger structures. Avascular tissue (tissue without blood vessels) can only grow so large before the interior becomes hypoxic, meaning oxygen-depleted. In avascular tumor spheroids, for example, the outer rim of cells receives oxygen and keeps dividing, while central cells become quiescent or die due to hypoxia. This produces a stratified structure with a necrotic core and a proliferative outer shell. Growth follows a Gompertz-like curve that plateaus rather than continuing indefinitely. The same principle applies to any growing tissue: mitosis can only sustain expansion where supply keeps up with demand.
Contact inhibition
When normal epithelial cells become densely packed and make contact with neighbors on all sides, they stop dividing. This is contact inhibition, and it is a fundamental brake on uncontrolled proliferation. The mechanism involves cadherin-cadherin junction signaling between adjacent cells, which dampens responsiveness to growth factor stimulation. Cells effectively sense crowding through mechanical and chemical signals and arrest mitosis in response. Research on epithelial monolayers has shown that this arrest becomes complete when individual cell area drops below a critical threshold, meaning physical space is literally the limiting variable.
Molecular brakes on division
Beyond physical constraints, specific molecular regulators actively suppress mitosis to prevent tissue overgrowth. Lkb1, for example, is a kinase that negatively regulates cell division in multiple tissue contexts. Tumor suppressor proteins, checkpoint kinases, and differentiation signals all contribute to a layered system of brakes. When these brakes fail, cells can divide past their normal boundaries, which is the basis of cancer. The fact that so many independent systems exist to stop mitosis tells you how seriously the body takes the risk of unchecked proliferation.
Quiescence and differentiation as deliberate stops
Adult stem cells in many tissues are held in a quiescent (non-dividing) state by signals from the surrounding cellular environment, sometimes called the niche. Cell-cell and cell-matrix interactions keep them inactive until injury or tissue need triggers activation. This is not a failure of the mitotic machinery. It is deliberate restraint. In wound healing, epithelial stem cells begin proliferating within roughly three days of injury and then return to a resting state as repair completes. Mitosis is turned on when needed, then turned off again.
What this means if you're trying to understand growth
The chain of logic here is worth summarizing cleanly. Mitosis produces two daughter cells from one. More daughter cells mean a higher total cell count. More cells arranged in organized layers and structures produce larger tissues. Larger tissues make larger organs. Larger organs make a larger organism. That is the direct line from a single mitotic event to whole-body growth. If you are asking how do organisms grow bigger at the whole-body level, it is essentially the same logic chain from more cells to larger organs to a larger organism.
But the process is never just 'divide until big.' It is a regulated, permission-based system where growth factors signal need, cyclin-CDK complexes grant access, checkpoints verify readiness, and physical and molecular limits set the ceiling. Understanding those constraints is just as important as understanding the mechanism of division itself, because they explain why organisms reach characteristic sizes and why losing those constraints causes disease.
If you want to go deeper on related questions, it is worth exploring how multicellular organisms coordinate growth across different tissue types, how single-celled organisms handle size limits differently without mitosis serving a growth function in the same way, and what the two main strategies for organismal growth actually are when you zoom out past just cell division. These two strategies explain what an organism can do to increase its size, either by adding cells or by increasing cell size two main strategies for organismal growth. Single-celled organisms grow too, but their size limits and growth strategies are different from mitosis-driven proliferation in multicellular development do single celled organisms grow. This is why the question of how big single-celled organisms can get depends on different size limits than those faced by mitosis-driven multicellular growth Single-celled organisms grow too.
FAQ
Does mitosis make every cell bigger so the whole organism grows?
Not always. Mitosis is a major driver of growth in tissues that are expanding, like embryos and healing wounds, but some cells grow mainly by increasing size or mass without dividing. Many neurons, for example, are largely post-mitotic, so tissue size changes there depend on other populations and on overall organ growth rather than repeated mitosis in the same cells.
Why don’t cells just get bigger each time they undergo mitosis?
A key reason is that the cell cycle requires a “window” for division, and daughters must recover their normal size during interphase. Right after cytokinesis, each daughter starts smaller than the parent, then grows in G1 before it can complete later steps and divide again.
If mitosis increases cell number, why isn’t growth simply proportional to division rate?
Cell number increases because each division roughly doubles a local cell population, but organism-level size also depends on coordinated growth of tissue structure (shape, spacing, and differentiation). If cells proliferate without correct patterning or mechanical organization, the tissue may grow abnormally rather than producing healthy, functional organs.
What happens to growth if mitosis keeps going but nutrient supply can’t keep up?
Developmental growth is limited by how quickly nutrients, oxygen, and growth signals can be delivered and used by the expanding tissue. Even if mitosis is active, larger tissues hit diffusion and supply limits, which can slow proliferation, push cells into quiescence, or trigger death in poorly supplied regions.
Is contact inhibition the same in all tissues, or only in epithelial layers?
Contact inhibition mainly matters in dense epithelial layers where cells physically crowd each other and reduce responsiveness to mitogenic cues. In other tissues with different architecture or in culture systems lacking proper cell-cell contact, proliferation can occur more readily because that crowding brake is weakened or absent.
Can an organism grow in size if its cells grow in mass but don’t divide much?
Yes, at least in principle, because cell growth and division can be regulated separately. In some contexts cells may increase mass and metabolism without dividing, while in others they may divide with altered growth timing (for example, shortening G1), which changes how average cell size evolves over time.
How do growth and cell death work together during development?
Most of the “net growth” comes from the balance of cell addition through division and cell removal through differentiation and programmed cell death. If proliferation increases but cell death also rises to the same extent, the tissue can stay the same size, so actual growth depends on the difference between those processes.
Why don’t all cells start dividing immediately when an organism needs more tissue?
In many tissues, stem cells are kept quiescent until a trigger in the niche says repair or maintenance is needed. That timing means mitosis is often localized to specific compartments, for example basal skin stem/progenitor layers, rather than happening everywhere at once.
How do mitosis checkpoints relate to cancer and abnormal growth?
Yes. If cells lose checkpoint control, DNA damage can be carried into division, increasing mutation load and potentially creating uncontrolled growth. Checkpoints like the G1 and G2/M gates exist to pause the cycle for repair or eliminate severely damaged cells, reducing the chance of tumor-like overgrowth.
If each division creates two cells, why do organisms eventually stop growing?
No, because cell doubling does not continue indefinitely due to molecular and physical limits. Constraints like diffusion limits, supply limits, mechanical space, and tumor-suppressive pathways create a ceiling, so growth typically slows and then plateaus even if cells can still divide.
What limits how long adult stem cells can keep proliferating?
Stem cell exhaustion or niche disruption can reduce the pool of competent dividing cells, leading to slower or halted tissue growth and regeneration. Likewise, differentiation programs can permanently remove cells from the mitotic pool, which helps tissues reach stable adult size.
Next Article
How Do Organisms Grow Bigger: Cells to Whole Bodies
How organisms grow bigger from cell division to organ scaling, plus limits like diffusion, energy, space, and regulation
